

Журнал высшей нервной деятельности им. И.П. Павлова, 2020, T. 70, № 2, стр. 231-242
СПОСОБНОСТИ К РАЗЛИЧЕНИЮ ВИДИМЫХ СИГНАЛОВ В ВОДНОМ ЛАБИРИНТЕ МОРРИСА У ВЫСОКО- И НИЗКОИМПУЛЬСИВНЫХ КРЫС
М. И. Зайченко 1, *, Г. Х. Мержанова 1, Г. А. Григорьян 1
1 Федеральное государственное бюджетное учреждение науки
Институт высшей нервной деятельности и нейрофизиологии Российской академии наук
Москва, Россия
* E-mail: mariya-zajchenko@yandex.ru
Поступила в редакцию 18.03.2019
После доработки 10.09.2019
Принята к публикации 16.09.2019
Аннотация
Высокоимпульсивные крысы быстрее, чем низкоимпульсивные обучались находить в водном лабиринте Морриса видимую платформу, на которой устанавливался специальный стимул-объект (флажок). У них также успешно вырабатывалась дифференцировка между парусом (дифференцировочный стимул-объект) и флажком, о чем можно было судить по достоверному уменьшению числа ошибочных заплывов в зону расположения паруса. В процессе дифференцировки ВИ крысы быстрее, чем НИ достигали платформы и проплывали при этом меньшее расстояние. В случае обратной дифференцировки (переделке) различия между группами по времени достижения платформы и пройденного расстояния исчезали. Но у ВИ крыс число заплывов к дифференцировочному объекту (теперь флажку) уменьшалось, чего не наблюдалось у НИ животных. Полученные данные свидетельствуют о том, что “эгоцентрические” задачи в водном лабиринте Морриса с выступающей платформой и флажком на ней легче решают высокоимпульсивные крысы, а “аллоцентрические” задачи, связанные с поиском скрытой под водой платформы, – низкоимпульсивные животные.
Импульсивность – это склонность субъекта к выполнению быстрых необдуманных действий. Она обнаруживается как одно из свойств индивидуально-типологических особенностей человека и животных. Чрезмерно выраженная импульсивность может сопровождать ряд серьезных психопатологий – шизофрению, маниакальные психозы, болезнь Паркинсона, агрессию, алкоголизм, наркоманию, ожирение, синдром дефицита внимания и гиперреактивности и др. [Benard et al., 2019; Canario et al., 2019; Linhartova et al., 2019; Vaughan et al., 2019; Brown et al., 2018; Johnson et al., 2017]. Выделяют двигательную импульсивность, импульсивность, связанную с расстройствами внимания и когнитивную импульсивность, зависимую от процессов принятия решения [Higgins, Silenieks, 2017; Зайченко и др., 2016; Cardinal et al., 2004; Logan et al., 1997; Mazur, 1987]. Для измерения и оценки каждого из перечисленных типов импульсивности существуют свои модели и приемы исследования [Eagle, Robbins, 2003; Robbins, 2002; Mazur, 1987]. Считается, что функциональная импульсивность связана с наиболее простым, экономным и не требующим особого напряжения когнитивных процессов поведением, если для этого имеется ситуация выбора [Salamone et al., 1994; Mazur, 1987]. Например, если предоставляется выбор между слабым немедленным и сильным задержанным подкреплениями, то импульсивный субъект (или животное) выбирает первую опцию [Mazur, 1987; Cardinal et al., 2004]. То же самое происходит при выборе слабого высоковероятного или сильного, но маловероятного подкрепления [Zoratto et al., 2016]. Если животному предоставляется выбор между получением легко добываемого слабого подкрепления и трудно добываемого сильного подкрепления, например, получения на одной стороне камеры подкрепления без препятствия, а на другой стороне с препятствием (переход через барьерную лесенку), то животные предпочитают первый вариант действия [Salamone et al., 1994]. В подобных случаях применимо даже такое понятие, как цена действия и выгода от него (cost-benefit). Причем всегда можно найти точку равновесия, при которой сложность выполнения задачи уравнивается величиной получаемой выгоды. Чем более импульсивным является субъект (или животное), тем больше точка равновесия сдвигается влево, т.е. имеет место предпочтение слабого и легко получаемого подкрепления. Недавно мы [Зайченко и др., 2016] показали, что у высокоимпульсивных крыс “сигнальная память” (cue memory) в 8-рукавном радиальном лабиринте проявляется лучше, чем у низкоимпульсивных животных. Высокоимпульсивные крысы обучались быстрее находить и поедать пищу в 4 подкрепляемых рукавах лабиринта со специальными вставками (дорожка из шерстяной ткани, металлическая сетка и т.д.), и при этом совершать меньше ошибок долгосрочной и краткосрочной памяти, чем низкоимпульсивные животные. Они также лучше осуществляли реакции избавления и избегания при собственном болевом электрокожном раздражении или при раздражении крысы партнера, издающего крик боли, по сравнению с низкоимпульсивными и амбивалентными животными (Зайченко и др., 2018). В работе [Зайченко и др., 2016а] в бассейне Морриса при поиске скрытой под водой платформы у высокоимпульсивных животных по ряду показателей (время нахождения платформы, пройденная дистанция и т.д.) краткосрочная и долгосрочная память проявлялись хуже, чем у низкоимпульсивных животных. Правда, различия касались суммарных эффектов Группы, но не эффектов взаимодействия Группы и Дней обучения. Из сказанного складывается впечатление, что животные с высокой степенью импульсивности легче решают задачи, связанные с выделением близлежащих стимулов обстановки (эгоцентрические), а животные с низкой степенью импульсивности – задачи, связанные с использованием отдаленных внешних стимулов (аллоцентрические) для “фиксации” местоположения скрытой в воде платформы или местоположения кормушек с подкреплением. Чтобы убедиться в справедливости этого тезиса, в настоящей работе мы провели специальную серию опытов на новых крысах в бассейне Морриса, но в отличие от предшествующей работы [Зайченко, 2016а] платформа не была скрыта под водой, а выдвигалась над ее поверхностью на 1.5–2.0 см. Дополнительно на платформе устанавливался флажок, который вместе с ее видимой частью сигнализировал о месте платформы в бассейне. Предполагалось, что если высокоимпульсивные крысы лучше, чем низкоимпульсивные животные решают эгоцентрические задачи, то они будут лучше и быстрее обучаться находить видимую платформу. В задачу работы входила проверка этого предположения. Дополнительно мы провели еще две серии опытов. В первой серии высоко- и низкоимпульсивные крысы обучались различать два видимых объекта, один из которых (флажок, положительный) сигнализировал истинное положение платформы, а другой (парус, отрицательный) – ее отсутствие. Во второй серии опытов проводили обратную дифференцировку (переделку) – парус становился теперь положительным стимулом, а флажок отрицательным.
МЕТОДИКА
Опыты проведены на 42 крысах-самцах линии Wistar в возрасте от 3 до 4 мес и массой 300–350 г. Крыс содержали в клетках группами по 4–5 животных при свободном доступе к воде. Количество пищи регулировалось так, чтобы поддерживать массу крысы на уровне приблизительно 80% от массы животного при свободном доступе к пище. Работа проводилась в два этапа. На первом этапе на 22 животных исследовали способность к обучению в бассейне Мориса на видимый сигнал. После анализа результатов, полученных на первом этапе, было принято решение повторить исследование на другой группе из 20 животных, добавив серии с дифференцировкой. Эксперименты осуществляли с соблюдением принципов гуманности, изложенных в директивах Европейского сообщества (2010/63/ЕU) и одобренных Комитетом по медицинской этике в соответствии с положением Института ВНД и НФ РАН о работе с экспериментальными животными.
Методика delay discounting. Вначале всех крыс обучали по этой методике для разделения на группы по степени импульсивности. Экспериментальная установка представляла собой прямоугольную камеру размером 30 × × 60 × 30 см. В середине передней стенки камеры была встроена кормушка, вход в которую загораживала подвижная шторка из прозрачного пластика. С двух сторон от кормушки размещались выдвижные педали. В кормушке и над каждой из педалей находились сигнальные лампочки. Управление включением ламп, выдвижением педалей, подача корма, формирование промежутков времени выполнялись с помощью персонального компьютера по программе, созданной Д.А. Ивлиевым. Крыс обучали нажимать на педаль для получения пищи – стандартной таблетки (dustless precision pellets) 45 мг производства фирмы “BioServ” (США). Обучение проводилось до тех пор, пока крыса с равной вероятностью не нажимала на каждую из педалей. Этот этап обучения длился 15–20 дней. Затем режим подкрепления меняли так, что нажатие на одну из педалей предоставляло 1 таблетку, тогда как нажатие на другую – 4 таблетки, но с задержкой в 5 с. Во время ежедневной экспериментальной сессии крысам предоставляли 25 проб. Схема, по которой проводилась каждая проба, была аналогична описанной в работах [Cardinal et al., 2002]. Проба начиналась с включением в камере лампочки общего освещения и одновременно лампочки в кормушке. В течение 10 с животному предоставлялась возможность отодвинуть носом шторку, закрывающую доступ в кормушку. При отсутствии такой реакции действие программы останавливалось, свет гас и через 5 с начинался новый цикл. Отодвигание шторки служило сигналом для выдвижения обеих педалей, которые оставались выдвинутыми в течение 10 с. При отсутствии реакции педали задвигались, свет гас и через 5 с производился перезапуск программы. При нажатии на одну из педалей крыса получала 1 таблетку сразу, на другую – 4 таблетки с задержкой. Связь педалей с задержкой, а также время задержки задавались в начале эксперимента. Таким образом, крысам предоставлялся выбор между получением слабого, но предоставляемого сразу после нажатия подкрепления (1 таблетка) и более ценного (4 таблетки), но отсроченного корма. Регистрировали число нажатий на обе педали и число пропусков реакций (k1 – число нажатий на педаль, предоставляющую слабое, но немедленное подкрепление, k2 – число нажатий на педаль для получения более ценного, но задержанного подкрепления. Рассчитывали долю задержанных реакций – отношение k2 к числу эффективных реакций k2/(k1 + k2). После введения задержки в получении пищи при нажатии на одну из педалей требовалось 10–15 сессий для стабилизации поведения по выбору подкрепления. Затем за следующие 10 сессий подсчитывали среднюю долю выбора задержанного подкрепления, на основании чего животное определяли в группы высокоимпульсивных (ВИ), низкоимпульсивных (НИ) и амбивалентных (АМБ).
Обучение в бассейне Морриса. Бассейн представлял собой круглую пластиковую емкость черного цвета диаметром 1.5 м и высотой 60 см (Noldus Ltd), наполненный водой до уровня 30 см. Он располагался в углу большой комнаты и был отделен от остального пространства темной матерчатой шторой. Бассейн условно, с помощью двух пересекающихся в центре прямых, разделяли на 4 равных квадранта. Кроме того, с помощью окружностей выделяли центральную, среднюю и периферическую зоны.
Точки пересечения прямых со стенками бассейна служили в качестве мест погружения животных в воду (условно – север, юг, запад, восток). В середину одного из квадрантов (средняя зона) помещали платформу (d10 см), верхний край которой был на 1.5–2.0 см выше уровня воды. Во время опытов в комнате было нормальное освещение 200 ± ± 30 люкс; температура воды в ходе опытов была 22–24°С.
Обучение находить платформу на видимый сигнал. В настоящей серии опытов крыс обучали находить безопасную платформу по зрительному объекту (флажку 10 × 5 см), фиксированному на поверхности платформы. Стержень, на котором крепился флажок, выступал на 10 см от поверхности платформы. Опыт продолжали 3 дня по 4 пробы в день с интервалом между пробами 10 мин. Каждую пробу начинали с погружения крыс в бассейн с разных его точек (север, юг и т.д.). Проба продолжалась до момента нахождения крысами безопасной платформы и залезания на нее, но не более 60 с. Если крыса не находила платформу в отведенное время, ее помещали туда принудительно и оставляли на платформе в течение 20 с. Положение платформы изменяли каждый день. Крыс разных групп брали в опыт в смешанном порядке, например, ВИ, НИ, АМБ, НИ и т.д.
Выработка дифференцировки двух видимых объектов, один из которых сигнализирует истинное местоположение платформы, а другой – ложное. Эту серию опытов продолжали 6 дней. Теперь на некотором расстоянии от видимой платформы, на которой по-прежнему помещался флажок, располагали еще один стимул-объект, близкий по размерам флажку (голубой парус), под которым платформы не было. Например, если платформа с флажком располагалась в квадранте 1, то парус был в квадранте 3. Крысы должны были научиться дифференцировать два стимула, один из которых показывает истинное положение безопасной платформы, а другой – ложное ее место. В день применяли 4 пробы. Максимальная продолжительность пробы составляла 60 с, интервалы между пробами – 10 мин.
Обратная дифференцировка. Переделка сигнального значения использовавшихся при исходном обучении стимул-объектов. На следующий день после завершения дифференцировки между флажком на платформе и парусом, ориентирующим животное к ее ложному местоположению, начинали выработку обратной дифференцировки. Теперь парус соответствовал истинному положению платформы, а флажок больше не был связан с ней и служил в качестве дифференцировочного стимул-объекта. Опыты продолжали по той же схеме, что и во время исходной дифференцировки, но поскольку крысы к этому времени очень быстро находили платформу, время переделки было сокращено до двух дней.
Поведение крыс в бассейне регистрировали с помощью системы видеорегистрации EthoVision 3.1 фирмы Noldus Ltd. Оценивали время с момента погружения крыс в бассейн до нахождения платформы и залезания на нее (латентный период), проплываемое до достижения платформы расстояние, число ошибок, т.е. число заплывов к дифференцировочному объекту, и скорость движения. Для каждой группы подсчитывали среднюю частоту ошибок, время достижения платформы и проплываемое до нее расстояние. Статистическую обработку проводили с помощью факторного анализа и анализа повторных измерений ANOVA с учетом межгруппового (Группа – ВИ/НИ/АМБ или ВИ/НИ) и внутригрупповых (Пробы, Дни обучения) факторов, используя программу STATISTICA 10. Средние значения групп сравнивали post hoc с помощью LSD теста Фишера.
РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЙ
Выделение крыс в группы с разной степенью импульсивности по методике delay discounting
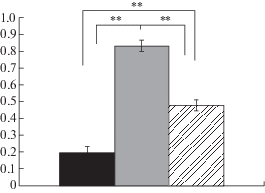
Период обучения крыс приоритетному выбору пищевого подкрепления разной ценности занимал 1–1.5 мес. В результате обучения были выделены три группы крыс, статистически значимо (p < 0.01) различающихся по проявлению задержанных реакций (рис. 1). Высокоимпульсивные животные, ВИ (n = 17) в процессе выбора между слабым немедленным и сильным задержанным подкреплением предпочитали выбирать первую комбинацию в более чем 60 процентах случаев. Низкоимпульсивные животные, НИ (n = 17) предпочитали выбор второй комбинации в более чем 60 процентах случаев. Амбивалентные животные, АМБ со смешанным типом реакций выбирали ту или другую комбинацию равновероятно.
Рис. 1.
Процент задержанных реакций у высокоимпульсивных, низкоимпульсивных и амбивалентных животных. По вертикали – доля задержанных реакций. Черным столбиком обозначены высокоимпульсивные животные, серым – низкоимпульсивные, и столбиком со штриховкой – амбивалентные крысы; ** – p < 0.01.
Fig. 1. The percentage of delayed reactions in high-, low impulsive and ambivalent animals. On vertical axis – a proportion of delayed reactions. Black columns – the high impulsive rats, grey columns – the low impulsive rats, shadowed columns – ambivalent rats. ** – p < 0.01.
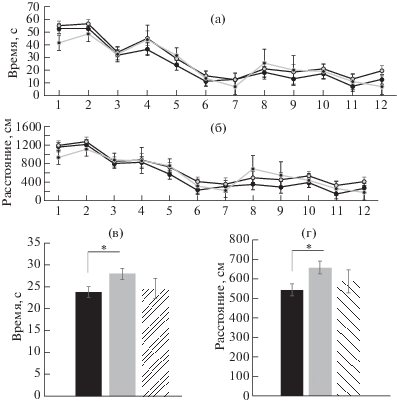
Динамика обучения высоко- и низкоимпульсивных крыс на видимый сигнал в водном лабиринте
Рис. 2 (а) демонстрирует динамику обучения крыс разных групп по латентному периоду достижения в лабиринте видимой платформы с флажком. Видно, что все группы крыс (ВИ, НИ и АМБ) быстро, в течение нескольких проб, находили платформу и залезали на нее. Латентный период достижения платформы за время обучения уменьшался с 55–40 с до 20–10 с. Дисперсионный факторный анализ (ANOVA) не обнаружил достоверного взаимодействия эффектов Группы и Проб [F(22, 380) = 0.36, p > 0.05], что свидетельствует об отсутствии различий в скорости обучения у крыс разных групп. То же самое обнаружилось при оценке динамики обучения крыс по показателю пройденного расстояния [F(22, 380) = 0.33, p > 0.05] (рис. 2 (б)). Между тем анализ суммарного эффекта Группы (для всех проб) выявил достоверные различия между группами по времени достижения платформы [F(2, 380) = 2.86, p < 0.05] за счет разницы между ВИ и НИ группами, p < 0.01) (рис. 2 (в)) и по пройденному расстоянию [F(2, 380) = 3.18, p < 0.05] за счет разницы между теми же группами, p < 0.01 (рис. 2 (г)). Эти результаты свидетельствуют о том, что хотя ВИ и НИ крысы примерно одинаково обучаются находить видимую в воде платформу, в целом (за все пробы) ВИ животные достигают ее быстрее и проплывают при этом меньше расстояния, чем НИ животные. Интересно, что скорость плавания у всех групп крыс мало отличалась друг от друга, судя по общему эффекту Группы [F(2, 380) = 1.41, p > 0.05] и по взаимодействию эффектов Группы и Проб [F(22, 380) = 0.85, p > 0.05].
Рис. 2.
Динамика нахождения крысами видимой платформы с флажком. (а) – время нахождения платформы; (б) – проплываемое расстояние. Сплошная черная линия указывает на динамику обучения у ВИ животных; пунктирная линия у НИ, а сплошная серая у амбивалентных крыс. По горизонтальной оси – номера проб; по вертикальной оси: на (а) и (в) – время нахождения платформы, на (б) и (г) – проплываемое расстояние. (в) – среднее суммарное (по всем пробам) время нахождения платформы; (г) – среднее суммарное проплываемое расстояние. * – p < 0.05.
Fig. 2. The dynamics of learning to reach a visible escape platform with bannerette. (а) – the latency to reach the platform; (б) – the distance swum. A solid black line indicates a dynamics of learning by the high impulsive animals; a solid gray line learning by the low impulsive animals and a dashed line – by the ambivalent rats. On horizontal axis – numbers of trials; on vertical axis: on (а) and (в) – the time of reaching the safe platform, on (б) and (г) – the distance swum. (в) – an averaged summed (by all trials) time of finding the safe platform. (г) – an averaged summed (by all trials) distance swum. * – p < 0.05.
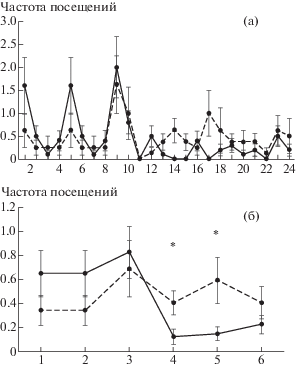
Дифференцировка высоко- и низкоимпульсивными крысами двух видимых объектов, один из которых сигнализирует истинное местоположение платформы, а другой – ложное
В данной и следующей серии экспериментов использовали 10 крыс ВИ группы, 8 крыс НИ группы и только 2 крысы амбивалентной группы (АМБ). В связи с малочисленностью последней группы мы не брали ее в анализ.
В экспериментах с дифференцировкой важным показателем является частота посещений зоны, равной 10 см вокруг объекта (парус), под которым не было платформы. На рис. 3 (а) показана частота посещения этого объекта крысами разных групп от 1-й до 24-й пробы. Следует отметить, что в первых пробах 1–3 дня ВИ крысы совершали больше ошибок, чем НИ животные, подплывая вначале к новому объекту, а не к платформе с флажком. У этой группы животных наблюдалось увеличение времени достижения платформы с 8.35 с ± 2.3 с в последней пробе предыдущей серии до 16.66 с ± 4.46 с в первой пробе дифференцировки, с разницей на уровне тенденции (р = 0.07). У них также увеличивалось проплываемое расстояние (р < 0.05) с 138 см ±23.17 см до 286.7 см ± 68.58 см. НИ крысы чаще подплывали к парусу на третий день дифференцировки, и в этот день у них тоже на уровне тенденции (р = 0.09) наблюдалось увеличение времени достижения платформы (с 11.08 с ± 2.4 с до 19.08 с ± 6.01 с), но изменение проплываемого расстояния (318 см ± 69.0 см и 401.47см ± 144.46 см) было недостоверным. У ВИ крыс по ходу обучения уменьшалось число посещений зоны дифференцировочного объекта (рис. 3 (б)) с достоверным эффектом Пробы [F(23, 216) = 1.6982, p < 0.05], что свидетельствует о выработке у них дифференцировки. Этого не происходило у НИ животных, которые хотя и продемонстрировали снижение частоты посещения объекта-паруса в последние дни обучения по сравнению с третьим днем, но оно было статистически не значимым. Взаимодействие эффектов Группы и Проб по частоте посещений дифференцировочного объекта (паруса) было также недостоверным [F(23, 384) = 1.1624, p = 0.28] но во второй половине этого этапа обучения крысы ВИ группы совершали статистически значимо меньше подходов к парусу с эффектом Группы [F(1, 210) = 12.225, p < 0.001].
Рис. 3.
Частота посещений дифференцировочного объекта. (а) – динамика изменений по пробам. По горизонтальной оси номера проб. (б) – динамика изменений по дням эксперимента. По горизонтальной оси номера дней эксперимента. По вертикальной оси число посещений. * – различия между группами в данный день эксперимента статистически значимы, p < 0.05. Остальные обозначения как на рис. 2.
Fig. 3. The frequency of disriminating object’s visits. (а) – the dynamics of changes by trials. On horizontal axis – numbers of trials. (б) – the dynamics of changes by days of experiments. On horizontal axis – numbers of days. * – the differences between the groups on this day of the experiment are statistically significant. The rest designations are the same as in Fig. 2.
На рис. 4 показаны время (а) достижения безопасной платформы и проплываемое до нее расстояние (б) в процессе дифференцировки. Надо сказать, что в первой серии опытов во время поиска видимой платформы с флажком крысы очень быстро обучались находить платформу. Так что ко второй серии опытов с дифференцировкой навык еще больше усовершенствовался и практически достиг нижнего асимптотического уровня (floor effect) с латентным периодом 10–12 с и проплываемым расстоянием 200–225 см. Несмотря на это, ВИ крысы проявили достаточно отчетливую динамику обучения [F(23, 216) = = 1.81, p < 0.05] в отличие от НИ животных. На последнем этапе ВИ крысы быстрее, чем НИ достигали безопасной платформы (рис. 4 (а)), хотя в последний день обе группы доплывали до платформы практически одинаково, несмотря на присутствие в бассейне дифференцировочного объекта. Различия между ВИ и НИ крысами проявились статистически значимо по взаимодействию эффектов Группы и Дней [F(5, 420) = 2.9164, p < 0.05] и по суммарному эффекту Группы во второй половине (4–6-е дни) дифференцировки [F(1, 210) =10.235, p < 0.01].
Рис. 4.
Динамика изменения времени достижения платформы (а) и проплываемого расстояния (б). По вертикальной оси время и расстояние соответственно, по горизонтальной – дни эксперимента. Остальные обозначения как на рис. 3.
Fig. 4. The dynamics of latency changes to reach a safe platform (а) and distance swum (б). On vertical axis – time and distance swum accordingly, on horizontal axis – days of experiments. The rest designations are the same as in Fig. 3.
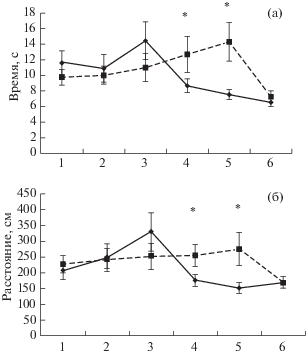
Примерно такая же картина наблюдалась при оценке пройденной дистанции у крыс ВИ и НИ групп (рис. 4 (б)). У ВИ крыс, в отличие от НИ животных, отмечалась отчетливая динамика обучения [F(23, 216) = 1.58, p < < 0.05]. Статистически значимые различия в суммарном эффекте Группы проявились также во втором периоде обучения [F(1, 210) = = 8.7459, p < 0.01].
Обратная дифференцировка. Переделка сигнального значения использовавшихся при исходном обучении стимул-объектов
В первой пробе во время выработки обратной дифференцировки в среднем по всем группам крыс наблюдалось увеличение времени достижения платформы по сравнению с последней пробой предыдущей серии (9.7 с ± ± 1.17 с и 7.1 с ± 0.6 с соответственно). Анализ повторных измерений ANOVA выявил статистически значимые различия между этими показателями [F(1, 16) = 4.7386, p < 0.05] (рис. 5 (а)). Проплываемое расстояние также увеличивалось с 163.2 см ± 18.7 см до 210.2 см ± ± 20.0 см, но различия в этом случае имели характер тенденции [F(1, 16) = 3.0855, p = 0.09]. Частота посещения объекта (флажка), ранее связанного с платформой, составляла 1.44 ± 0.2, что свидетельствует о сохранившейся связи между этим объектом и безопасной платформой.
Рис. 5.
Переделка оригинальной дифференцировки. (а) – различия по времени нахождения платформы между последней пробой серии с дифференцировкой (светлый столбик) и первой пробой серии с переделкой (темный столбик) суммарно, по всем группам крыс. (б) – динамика изменения проплываемого расстояния и (в) – числа посещений дифференцировочного объекта ВИ и НИ группами крыс. По горизонтальной оси – номера проб. Остальные обозначения как на рис. 3.
Fig. 5. Reversal of discrimination learning. (а) – differences in times of reaching a safe platform in the last trial of original learning (white column) and the first trial of reversal learning (dark columns) in total by all groups of rats. (б) – the dynamics of distance swum over the trials, and (в) – the same for the dynamics of a number of discriminating object visits by HI and LI rats. On horizontal axis – numbers of trials. The rest designations are the same as in Fig. 3.
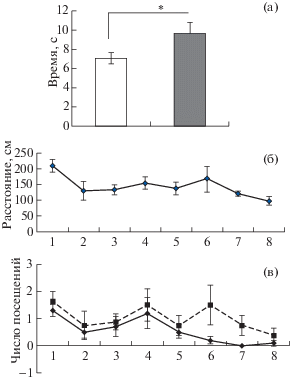
На протяжении семи последующих проб наблюдались следующие изменения. При оценке проплытого расстояния анализ фактора Пробы показал статистически значимое его уменьшение [F(7, 128) = 2.2363, p < 0.05]. На рис. 5 (б) показана динамика изменения расстояния по ходу выработки переделки. Также была выявлена тенденция в снижении времени достижения платформы [F(7, 128) = = 1.8978, p = 0.075]. Все это свидетельствует о том, что обучение при переделке происходило, но как видно из рисунков в основном в ВИ группе. В то же время статистически значимых различий между ВИ и НИ животными по фактору Группа для пройденного расстояния [F(1, 128) = 2.1694, p > 0.05] и для времени достижения платформы [F(1, 128) = 1.22, p > > 0.05] не наблюдалось. Не было также достоверных различий по эффектам взаимодействия Группы и Проб [F(7, 128) = 0.8667, p > > 0.05] [F(7, 128) = 0.4384, p > 0.05] для пройденной дистанции и времени достижения платформы соответственно.
При анализе частоты посещений зоны флажка (бывшего положительного сигнального объекта при оригинальной дифференцировке) отчетливая динамика обучения, судя по эффекту Пробы [F(7, 72) = 2. 942, p < 0.01], наблюдалась только у крыс ВИ группы. У НИ крыс она не проявилась [F(7, 56) = 0.9768, p = = 0.457]. ВИ крысы реже посещали дифференцировочный объект, о чем свидетельствовал анализ фактора Группы [F(1, 128) = 6.0511, p < 0.05]. В то же время анализ взаимодействия Группы и Проб статистически значимых отличий не выявил [F(7, 128) = 0.5456, p > 0.05] (рис. 5 (в)).
ОБСУЖДЕНИЕ РЕЗУЛЬТАТОВ
В настоящих опытах ВИ животные быстрее находили видимую платформу с флажком, и проплывали до нее меньшее расстояние, чем НИ животные. Они также лучше дифференцировали объекты, связанные с истинным и ложным местоположением безопасной платформы. Это проявилось в меньшей частоте посещений ВИ крысами зоны (10 см) дифференцировочного объекта (паруса). Причем только у ВИ крыс наблюдалось достоверное уменьшение числа ошибочных заплывов к зоне дифференцировочного объекта в ходе обучения, что говорит о выработке у них дифференцировки на флажок и парус. ВИ животные обучались также переделке сигнального значения исходных стимул-объектов, используемых при оригинальной дифференцировке, судя по уменьшению числа ошибочных заплывов к новому дифференцировочному стимул-объекту (флажку). Полученные результаты свидетельствуют о том, что ВИ крысы способны лучше, чем НИ решать эгоцентрические задачи, связанные с внутрилабиринтными стимулами в водном лабиринте Морриса. При усложнении задачи, например, в случае поиска скрытой под водой платформы ВИ решают эти задачи хуже, чем НИ крысы [Зайченко и др., 2016 б].
Функциональную (естественную) импульсивность связывают с расстройствами механизмов внимания, нарушением когнитивного контроля и неспособностью к торможению двигательных реакций [Patros et al., 2016; Mitchell et al., 2015; Robbins, 2002; Logan et al., 1997]. На сегодняшний день все исследователи склоняются к тому, что импульсивность связана со снижением внимания и чрезмерной двигательной активностью, что находит отражение в хорошо известном синдроме дефицита внимания и гиперреактивности у детей и взрослых [Patros et al., 2016]. Т. Роббинсом [Robbins, 2002] была разработана специальная методика дифференцировки по месту (5-choice serial reaction time task, CSRTT) [Carli et al., 1983; Blondel et al., 1999], которая с разными модификациями на протяжении многих лет используется в качестве основного приема для исследования внимания при импульсивном поведении. Животному предоставляется выбор в ответ на включение условного раздражителя (вспышки света, 5 с) в одном из 5 окошек получать подкрепление. Оценивают ошибки выбора, преждевременные реакции в межсигнальный период и пропуски реакций. В этой модели ВИ животные совершают намного больше ошибок и преждевременных реакций, чем НИ. Нетрудно видеть, что оценка внимания по различению стимулов, сигнализирующих истинное и ложное местоположение платформы в водном лабиринте Морриса, в наших опытах существенно отличается от общепринятой оценки по методу 5-CSRTT. Главным отличием, на наш взгляд, является степень сложности решения задачи. В частности, в модели 5-CSRTT вероятность правильного выбора составляет 20%, в то время как в использованной нами модели она составляет 100%, поскольку при совершении ошибки проба не заканчивается, а поведение ориентируется на новый поиск безопасной платформы. В модели 5-CSRTT время принятия решения и действия составляет всего 5 с. Если действие за это время не выполняется, то наступает пропуск реакции, а если оно выполняется преждевременно, то засчитывается как ошибочное и проба заканчивается. В наших опытах в водном лабиринте максимальное затратное время на дифференцировку составляет 60 с, хотя в реальности крысы решают эту задачу намного быстрее, обычно за 10–15 с или меньше. Следует учитывать также фактор, мотивирующий поведение. Одно дело выполнять задачу, связанную с получением положительного подкрепления (как в модели 5-CSRTT), а другое – совершать действия, направленные на устранение опасности (поиск безопасной платформы). О том, что дифференцировка внутренних лабиринтных стимулов является не очень сложной задачей, демонстрируют опыты Л. Робинсона и соавт. [Robinson et al., 2001], в которых животные легко обучались дифференцировать две карточки, состоящие из вертикальных черных и белых полосок (шириной от 1.0 до 10.0 см) с визуальной точностью 1.5 с/deg. При переделке предшествующего навыка на новый, в котором ранее положительный условный стимул сигнализирует теперь ее ложное положение (отсутствие платформы), а бывший отрицательный условный стимул – реальное положение платформы, задача существенно усложняется [Izquierdo, Jentsch, 2012] за счет конфликта между старым и новым опытом. В этой ситуации различия в поведении у ВИ и НИ животных при поиске платформы практически исчезают. Следует отметить, что у ВИ животных наблюдалось постепенное снижение числа подходов к дифференцировочному объекту, что проявилось в достоверном уменьшении числа ошибок в ходе обучения. В то же время НИ крысы в первые пробы обучения с дифференцировкой реже подплывали к новому объекту (парусу). Вероятно, это объясняется более высоким уровнем тревожности этой группы крыс и их более выраженной реакцией на новизну, что было показано в нашей предыдущей работе [Левандовская, Зайченко, 2013].
В литературе имеются всего несколько работ, свидетельствующих о прямой связи импульсивности с рабочей и долгосрочной памятью у крыс [Зайченко и др. 2016 а, б; Renda et al., 2014; Dellu-Hagedorn, 2006]. В этих работах вначале с помощью методики “delay discounting” выделяли группы животных с высокой и низкой степенью импульсивности, а затем исследовали у них состояние памяти (пространственной и сигнальной, рабочей и долгосрочной) в моделях обучения с использованием лабиринтов или дифференцировок типа “выбор в зависимости от местоположения” (matching to position task). Было показано, что рабочая память в 8-рукавном радиальном [Dellu-Hagedorn, 2006] и водном лабиринте Морриса [Зайченко и др., 2016 б] у ВИ и НИ крыс мало отличается друг от друга, хотя в работе [Зайченко и др., 2016 б] ВИ крысы решали пространственную задачу в целом хуже, чем НИ животные. В другой работе [Renda et al., 2014] в процессе выработки дифференцировки “выбор в зависимости от местоположения” рабочая память была хуже выражена у импульсивных животных. Следует, однако, иметь в виду, что авторы использовали для этого довольно трудную задачу. Она начиналась десятикратным нажатием крысой на одну из педалей (образец), которая располагалась на передней стенке камеры. Затем крыса должна была перейти к педали, расположенной на задней стенке, нажатие на которую после определенной задержки открывало доступ к двум педалям на передней стенке. Нажатие на педаль-образец считалось правильной реакцией, а нажатие на другую педаль – ошибкой [Renda et al, 2014]. При решении более простой задачи, а именно, нахождении пищи в 4 рукавах 8-рукавного радиального лабиринта со специальными вставками в подкрепляемых рукавах, у ВИ животных рабочая и долгосрочная память проявлялись лучше, чем у НИ крыс [Зайченко и др., 2016 а]. Они быстрее обходили все подкрепляемые рукава лабиринта и делали при этом меньше ошибок, чем НИ животные. В ряде работ пространственную и сигнальную память исследовали у крыс с искусственно созданной импульсивностью, которую вызывали с помощью вмешательств в раннем постнатальном периоде [Bañuelos et al., 2012], или путем повреждения некоторых структур мозга [Mariano et al., 2009; Cheung, Cardinal, 2005], или у спонтанно гипертензивных крыс (spontaneously hypertensive rats), обладающих высокоимпульсивными свойствами [Ferguson, Cada, 2004]. В частности, С. Бануэлош и соавт. [Bañuelos et al., 2012] на 4–9-й дни постнатальной жизни вводили крысятам этиловый спирт (5.25 г/кг в день) и на 35–38-й дни тестировали у них степень импульсивности и поведение в водном лабиринте Морриса. Крысы, получавшие спирт, с одной стороны, проявляли большую степень импульсивности, а с другой, так же легко, как и контрольные животные, находили видимую платформу, выделенную специальными стимулами. Однако более сложную пространственную задачу (поиск платформы под водой) они решали хуже, чем контрольные животные. Спонтанно гипертензивные ВИ крысы лучше обучались в водном лабиринте Морриса [Ferguson, Cada, 2004], чем контрольные (Wistar-Kyoto) животные. В другой работе [Sontag et al., 2013] спонтанно гипертензивные ВИ крысы в водном лабиринте Морриса проявляли такую же пространственную рабочую и долгосрочную память, что и контрольные животные, но с учетом более выраженной у них двигательной активности они решали задачи в целом лучше, чем контрольные крысы. Эти результаты совпадают с нашими данными [Зайченко и др., 2016 б] по исследованию пространственной рабочей и долгосрочной памяти у крыс, выделенных в ВИ и НИ группы с помощью методики delay-discounting. Известно, что повреждения орбито-фронтальной коры и гиппокампа вызывают у крыс импульсивное поведение, которое оценивали по методике delay discounting [Mariano et al., 2009; Cheung, Cardinal, 2005]. Причем крысы с повреждениями орбито-фронтальной коры решали сложные пространственные задачи в водном лабиринте хуже, чем контрольные животные, а задачи, связанные с различением черно-белых и серых полосок в Т-образном лабиринте примерно так же, как и контрольные крысы [Mariano et al., 2009]. В работе [Schoenbaum et al., 2002] было показано, что крысы с повреждениями орбито-фронтальной коры дифференцируют запахи, запускающие go/no go реакции, так же, как и контрольные крысы, но с усложнением задачи, в частности, при выработке обратной дифференцировки, они справлялись с ее решением хуже, чем контрольные животные.
Таким образом, все сказанное свидетельствует о том, что ВИ животные лучше, чем НИ крысы решают эгоцентрические задачи, связанные с использованием внутрилабиринтных стимулов и сигнальной памяти (cue memory), и хуже аллоцентрические задачи, связанные с использованием внелабиринтных стимулов и пространственной гиппокамп-зависимой памяти.
ВЫВОДЫ
1. Высокоимпульсивные крысы быстрее, чем низкоимпульсивные обучались находить в водном лабиринте Морриса видимую платформу, на которой устанавливался специальный стимул-объект (флажок), и проплывали до нее меньшее расстояние.
2. Высокоимпульсивные крысы успешно вырабатывали дифференцировку между парусом (дифференцировочный стимул-объект) и флажком (сигнальный стимул-объект), о чем можно было судить по достоверному уменьшению числа ошибочных заплывов в зону расположения паруса. В процессе дифференцировки ВИ крысы быстрее, чем НИ достигали платформы и проплывали при этом меньшее расстояние.
3. В случае обратной дифференцировки (переделки) различия между группами по времени достижения платформы и пройденного расстояния исчезали. Но у ВИ крыс число ошибочных заплывов к дифференцировочному объекту (флажку) уменьшалось, чего не наблюдалось у НИ животных.
4. Приведенные результаты вместе с нашими ранними данными свидетельствуют о том, что ВИ крысы лучше, чем НИ животные решают эгоцентрические задачи, связанные с использованием внутрилабиринтных зрительных стимулов, и хуже аллоцентрические задачи, связанные с использованием внелабиринтных пространственных стимулов.
Список литературы
Левандовская А.А., Зайченко М.И., Мержанова Г.Х. Оценка исследовательской активности и тревожности крыс, различающихся по выраженности импульсивного поведения. Журн. высш. нервн. деят. им. И.П. Павлова. 2013. 63 (6): 719–729.
Зайченко М.И., Григорьян Г.А., Мержанова Г.Х. Реакции избавления и избегания на собственное и чужое болевое электрокожное раздражение у крыс с высокой и низкой степенью импульсивности. Журн. высш. нервн. деят. им. И.П. Павлова. 2018. 68 (4): 477–487.
Зайченко М.И., Шаркова А.В., Григорьян Г.А., Мержанова Г.Х. Импульсивность улучшает сигнальную память в 8-канальном лабиринте у крыс. Журн. высш. нервн. деят. им. И.П. Павлова. 2016 а. 66 (5): 600–610.
Зайченко М.И., Баженова Д.А., Григорьян Г.А., Мержанова Г.Х. Влияет ли свойство импульсивности на проявления долгосрочной и краткосрочной памяти у крыс? Журн. высш. нервн. деят. им. И.П. Павлова. 2016 б. 66 (1): 82–91.
Bañuelos C., Gilbert R.J., Montgomery K.S., Fincher A.S., Wang H., Frye G.D., Setlow B., Bizon J.L. Altered spatial learning and delay discounting in a rat model of human third trimester binge ethanol exposure. Behav Pharmacol. 2012. 23 (1): 54–65.
Bénard M., Bellisle F., Kesse-Guyot E., Julia C., Andreeva V.A., Etilé F., Reach G., Dechelotte P., Tavolacci M.P., Hercberg S., Péneau S. Impulsivity is associated with food intake, snacking, and eating disorders in a general population. Am. J. Clin. Nutr. 2019.109 (1):117–126.
Blondel A., Simon H., Sanger D.J., Moser P. The effect of repeated nicotine administration on the performance of drug-naive rats in a five-choice serial reaction time task. Behav Pharmacol. 1999. 10 (6–7): 665–673.
Brown H.E., Hart K.L., Snapper L.A., Roffman J.L., Perlis R.H. Impairment in delay discounting in schizophrenia and schizoaffective disorder but not primary mood disorders. N.P.J. Schizophr. 2018. 4 (1): 9.
Canário N., Sousa M., Moreira F., Duarte I.C., Oliveira F., Januário C., Castelo-Branco M. Impulsivity across reactive, proactive and cognitive domains in Parkinson’s disease on dopaminergic medication: Evidence for multiple domain impairment. PLoS One. 2019. 14 (2): e0210880.
Cardinal R.N., Winstanley C.A., Robbins T.W., Everitt B.J. Limbic corticostriatal systems and delayed reinforcement. Ann N. Y. Acad Sci. 2004. 1021: 33–50.
Carli M., Robbins T.W., Evenden J.L., Everitt B.J. Effects of lesions to ascending noradrenergic neurones on performance of a 5-choice serial reaction task in rats; implications for theories of dorsal noradrenergic bundle function based on selective attention and arousal. Behav Brain Res.1983.9 (3): 361–80.
Cheung T.H., Cardinal R.N. Hippocampal lesions facilitate instrumental learning with delayed reinforcement but induce impulsive choice in rats. BMC Neurosci. 2005. 13 (6): 36.
Dellu-Hagedorn F. Relationship between impulsivity, hyperactivity and working memory: A differential analysis in the rat. Behav. Brain Funct. 2006. 2: 10. https://doi.org/10.1186/1744-9081-2-10
Eagle D.M., Robbins T.W. “Inhibitory control in rats performing a stop-signal reaction-time task: effects of lesions of the medial striatum and d-amphetamine,” Behav. Neurosci., Psychopharmacology. 2003. 117 (6): 1302–1317.
Ferguson S.A., Cada A.M. Spatial learning/memory and social and nonsocial behaviors in the spontaneously hypertensive, Wistar-Kyoto and Sprague-Dawley rat strains. Pharmacol Biochem Behav. 2004.77 (3): 583–594.
Higgins G.A., Silenieks L.B. Rodent test of attention task and impulsivity: the 5-choice serial reaction time. Curr. Protoc. Pharmacol. 2017. 78: 5.49.1–5.49.34.
Johnson S.L., Carver C.S., Tharp J.A. Suicidality in Bipolar Disorder: The Role of Emotion-Triggered Impulsivity. Suicide Life Threat Behav. 2017. 47 (2): 177–192.
Izquierdo A., Jentsch J.D. Reversal learning as a measure of impulsive and compulsive behavior in addictions.Psychopharmacology. 2012.219 (2): 607–620.
Linhartová P., Širůček J., Ejova A., Barteček R., Theiner P., Kašpárek T. Dimensions of Impulsivity in Healthy People, Patients with Borderline Personality Disorder, and Patients with Attention-Deficit/Hyperactivity Disorder. J. Atten. Disord. 2019.an 10:1087054718822121.
Logan G.D., Schachar R.J., Tannock R. Impulsivity and Inhibitory Control Psychological Science. 1997 (1): 60–64.
Mariano T.Y., Bannerman D.M., McHugh S.B., Preston T.J., Rudebeck P.H., Rudebeck S.R., Rawlins J.N., Walton M.E., Rushworth M.F., Baxter M.G., Campbell T.G. Impulsive choice in hippocampal but not orbitofrontal cortex-le-sioned rats on a nonspatial decision-making maze task. Eur. J. Neurosci. 2009. 3: 472–484.
Mazur J. An adjusting procedure for studying delayed reinforcement. In Quantitative Analysis of Behaviour. The Effect of Delay and Intervening Events on Reinforcement Value, M.L. Commons, J.A. Nevin, and H.C. Rachlin (eds.), Erlbaum, Hillsdale, N. J. 1987. 5: 55–73.
Mitchell S.H., Napier T.C., Reynolds B., Schütz C.G., Setlow B., Sher K.J., Swann A.C., Tedford S.E. White M.J., Winstanley C.A., Yi R., Potenza M.N., Moeller F.G. Choice impulsivity: Definitions, measurement issues, and clinical implications. Personal Disord. 2015. 6 (2): 182–198.
Patros C.H., Alderson R.M., Kasper L.J., Tarle S.J., Lea S.E., Hudec K.L. Choice-impulsivity in children and adolescents with attention deficit/hyperactivity disorder (ADHD): A meta-analytic review. Clin. Psychol. Rev. 2016. 43: 162–174.
Renda C.R., Stein J.S., Madden G.J. Impulsive choice pre-dicts poor working memory in male rats. PloS One. 2014. 9 (4): e93263.
Robbins T.W. The 5-choice serial reaction time task: behavioral pharmacology and functional neurochemistry. Psychopharmacology2002, 163 (3–4): 362–380.
Robinson L., Bridge H., Riedel G. Visual discrimination learning in the water maze: a novel test for visual acuity. Behav. Brain Res. 2001. 119 (1): 77–84.
Salamone J.D., Cousins M.S., Bucher S. Anhedonia or anergia? Effects of haloperidol and nucleus accumbens dopamine depletion on instrumental response selection in a T-maze cost/benefit procedure. Behav Brain Res. 1994. 5 (2): 221–229.
Schoenbaum G., Nugent S.L, Saddoris M.P., Setlow B. Orbitofrontal lesions in rats impair reversal but not acquisition of go, no-go odor discriminations. Neuroreport. 2002. 13 (6): 885–890.
Sontag T.A., Fuermaier A.B., Hauser J., Kaunzinger I., Tucha O., Lange K.W. Spatial memory in spontaneously hypertensive rats (SHR). PLoS One. 2013. 8 (8): e74660.
Vaughan C.L., Stangl B.L., Schwandt M.L., Corey K.M., Hendershot C.S., Ramchandani V.A. The relationship between impaired control, impulsivity, and alcohol self-administration in nondependent drinkers. Exp. Clin. Psychopharmacol. 2019. https://doi.org/10.1037/pha0000247
Zoratto F., Laviola G., Adriani W. The subjective value of probabilistic outcomes: Impact of reward magnitude on choice with uncertain rewards in rats. Neurosci Lett. 2016. 617: 225–231.
Дополнительные материалы отсутствуют.
Инструменты
Журнал высшей нервной деятельности им. И.П. Павлова