

Журнал высшей нервной деятельности им. И.П. Павлова, 2019, T. 69, № 5, стр. 570-576
ПОЛИМОРФИЗМ STIN2VNTR ГЕНА ТРАНСПОРТЕРА СЕРОТОНИНА: АССОЦИАЦИИ С ЭФФЕКТИВНОСТЬЮ КРАТКОВРЕМЕННОЙ ПАМЯТИ У МОЛОДЫХ И ПОЖИЛЫХ ИСПЫТУЕМЫХ
Н. В. Вольф 1, 2, *, Е. Ю. Приводнова 1, 2, Д. В. Базовкина 1, 2, 3
1 Научно-исследовательский институт физиологии и фундаментальной медицины
Новосибирск, Россия
2 Новосибирский государственный университет
Новосибирск, Россия
3 Институт цитологии и генетики СО РАН
Новосибирск, Россия
* E-mail: volf@physiol.ru
Поступила в редакцию 14.06.2018
После доработки 15.01.2019
Принята к публикации 11.02.2019
Аннотация
Показано, что когнитивные способности и скорость их угасания с возрастом в значительной степени определяются генетическими факторами. Серотонинергическая система мозга влияет на развитие, пластические перестройки мозга и играет важную роль в процессах обучения и памяти. Обнаруженная связь активности гена транспортера серотонина с эффективностью процессов памяти послужила основой для изучения ассоциаций между эффективностью памяти и полиморфизмом STin2VNTR, который обусловлен изменением числа тандемных повторов во втором интроне с двумя частыми (10 и 12 повторов) и одним редким (9 повторов) аллелями. Этот полиморфизм выполняет роль аллель-зависимого усилителя экспрессии гена. В исследовании принимали участие молодые (n = 204: 84 мужчины и 120 женщин, 19–35 лет) и пожилые (n = 129: 57 мужчин и 72 женщины, старше 55 лет) испытуемые-правши. Эффективность вербальной памяти определяли по воспроизведению слов c правого и левого уха в процедуре дихотического тестирования, а также по показателям распознавания среди дистракторов, ранее предъявленных на экране монитора слогов; образной – по распознаванию фигур. Независимо от других факторов носители генотипа 12/12 полиморфизма STin2VNTR, по сравнению с носителями аллеля 10, демонстрировали достоверно лучшие показатели запоминания дихотически предъявлявшихся слов и образной памяти. Для образной памяти эффект генотипа был более выражен в старшей возрастной группе.
С увеличением продолжительности жизни особое значение приобретает проблема сохранения умственного потенциала при старении. Показано, что когнитивные способности и скорость их угасания с возрастом в значительной степени определяются генетическими факторами, связанными с полиморфизмом в генах серотонинергической системы [Payton et al., 2005].
Серотонинергическая система мозга влияет на развитие, пластические перестройки мозга и играет важную роль в процессах обучения и памяти [Sodhi, Sanders-Bush, 2004; Homberg, et al., 2014; van Kesteren, Spencer 2003; Meneses, 1999]. Одним из ключевых регуляторов функций этой системы является транспортер серотонина (5-HTT), осуществляющий обратный захват медиатора из синаптической щели. Изучение гена SLC6A4, кодирующего белок транспортер серотонина, выявило два функциональных полиморфизма. Первоначально был найден полиморфный участок 5-HTTLPR в промоторной области гена, состоящий из 16 (long, L) или 14 (short, S) повторяющихся элементов. Второй полиморфизм, STin2VNTR, выявлен позже и обусловлен изменением числа тандемных повторов во втором интроне с двумя частыми (10 и 12 повторов) и одним редким (9 повторов) аллелями. Этот полиморфизм выполняет роль аллель-зависимого усилителя экспрессии гена, то есть усиливающий эффект полиморфизма зависит от количества повторов в аллеле, и является разным для аллелей 9, 10 и 12 [Fiskerstrand et al., 1999]. Влияние полиморфизмов гена 5-HTT человека широко изучается в отношении регуляции аффективных состояний и расстройств. Однако при изучении ассоциации 5-HTT с эффективностью когнитивных функций у здоровых испытуемых, в том числе и при непатологическом старении, изучался почти исключительно полиморфизм 5-HTTLPR [Пучкова, Дорохов, 2015; Borg et al., 2009; Madsen et al., 2011; Roiser et al., 2007; Вольф и др., 2017].
Настоящее исследование направлено на выявление ассоциаций полиморфизма STin2VNTR гена транспортера серотонина и эффективности кратковременной памяти у здоровых испытуемых молодого и пожилого возраста.
МЕТОДИКА
Испытуемые. В исследовании принимали участие молодые (n = 204: 84 мужчины и 120 женщин, 19–35 лет) и пожилые (n = 129: 57 мужчин и 72 женщины, старше 55 лет) испытуемые правши. Все испытуемые учились или работали на момент исследования. Это были студенты и сотрудники высших учебных заведений г. Новосибирска и научно-исследовательских институтов Сибирского отделения РАН, а также лица, привлеченные по объявлениям. Участники исследования имели нормальное или скорректированное до нормального зрение, не имели психических нарушений, травматических или иных поражений головного мозга. Не допускались лица с перенесенными инфарктами и диабетом. Из эксперимента с дихотическим тестированием были исключены испытуемые с ослаблением слуха (8 человек старшей возрастной группы (СВГ): 3 мужчины и 5 женщин). Все испытуемые дали информированное согласие на участие в исследовании. Исследование было одобрено Этическим комитетом Института физиологии и фундаментальной медицины.
Дихотическое тестирование. Синхронно по одному слову в каждое ухо со скоростью 1 пара в секунду предъявляли 10 пар одно- или двухсложных существительных с инструкцией распределения внимания на оба уха. Использовали 7 списков слов, после каждого из них следовал период (1 мин) свободного письменного воспроизведения запомненных слов. Определяли число запомненных слов, подававшихся в левое ухо (адресованных правому) и правое ухо (адресованных левому полушарию).
Определение эффективности вербальной и образной памяти. В исследовании применяли компьютеризированную методику: в одной серии последовательно в центре монитора предъявляли для запоминания 10 трехбуквенных слогов, в другой – 10 простых геометрических фигур (время предъявления 1 с, интервал между стимулами 1 с). В каждой серии сразу после запоминания следовало тестирование с предъявлением случайной последовательности из 20 слогов или 20 фигур, среди которых 10 не предъявлялись ранее для запоминания. Испытуемые должны были нажимать на клавиши “да” и “нет” при появлении на экране соответственно знакомого или нового стимула. Регистрировали число ошибок для слогов и фигур.
Генетическое исследование. Клетки для анализа ДНК получали из соскоба слизистой с внутренней поверхности щеки. ДНК выделяли солевым методом [Miller et al., 1988]. Для этого клетки в течение двух часов лизировали при 50°C с помощью протеазы К. Белки осаждали насыщенным раствором NaCl (6M). Отбирали супернатант, затем ДНК осаждали с помощью 2 объемов 96% этанола, отмывали 70% этанолом, высушивали, растворяли в стерильной воде и хранили до использования при температуре –20°С. Генотипы 9/9, 9/10, 9/12, 10/10, 10/12 и 12/12 полиморфизма STin2VNTR, локализованного во втором интроне гена транспортера серотонина, определяли с помощью ПЦР реакции со специфическими к данному полиморфизму праймерами 5'-GGTCAGTATCACAGGCTGCGAGTAG-3' и 5'-TGTTCCTAGTCTTACGCCAGTGAAG-3' (температура отжига 63°С). Последующее разделение и визуализация продуктов ПЦР осуществлялись методом электрофореза на 3% агарозном геле с окрашиванием бромистым этидием для визуализации. Размеры фрагментов STin2.12, STin2.10 и STin2.9 аллелей были 298, 265 и 248 п.о. соответственно [Avula et al., 2011]. Учитывая редкую встречаемость аллеля 9, мы объединили аллель 9 в одну группу с аллелем 10, более редким по сравнению с аллелем 12. Таким образом, генотипирование сводят к 3-аллельному варианту [Fan, Sklar, 2005], рассматривая деление на группы (12/12), (12/10 и 12/9) и (10/10 и 10/9), обозначенные нами соответственно как 12/12, 12/10' и 10'/10', чтобы указать, что это не “чистые” генотипы 10/10 и 10/12, а и генотипы, включающие и аллель 9. Статистический анализ проводили с использованием repeated measures ANOVA по программе STATISTICA 7. Последующий анализ проводили с помощью метода плановых сравнений и критерия Фишера.
РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЙ
Распределение генотипов, c учетом трехаллельного полиморфизма в младшей (Chi2 = 5.91, p = 0.41) и старшей (Chi2 = 1.48, p = 0.35) возрастных группах, не отличалось от соотношения Харди-Вайнберга. Частоты для полиморфизма STin2VNTR гена транспортера серотонина приведены в табл. 1. По частотам представленных генотипов не было различий между группами испытуемых разного возраста (Chi2 = 1.79, p = 0.41).
Таблица 1.
Частоты для объединенного по аллелям 9 и 10 и триаллельного полиморфизма STin2VNTR у молодых и пожилых испытуемых Table 1. Frequencies for joint (9 with 10 alleles) and for triallelic STin2VNTR polymorphism in young and elder participants
| Молодые испытуемые | Испытуемые старшей группы | |||||
|---|---|---|---|---|---|---|
| Объединенные (%) | 10'/10' (22.0) | 12/12 (37.3) | 10'/12 (40.7) | 10'/10' (16.2) | 12/12 (41.9) | 10'/12' (41.9) |
| и триаллельные генотипы (N) | 10/10 (42) | 12/12 (76) | 10/12 (81) | 10/10 (19) | 12/12 (54) | 10/12 (51) |
| 9/10 (3) | 9/12 (2) | 9/10 (2) | 9/12 (3) | |||
| 9/9 (0) | 9/9 (0) | |||||
| Частоты аллелей | 9 (5) | 10(168) | 12(235) | 9(5) | 10(91) | 12(162) |
Дисперсионный анализ ассоциаций полиморфизма Stin2VNTR гена транспортера серотонина и эффективности воспроизведения слов, адресованных левому и правому полушариям, в дихотическом тесте проведен с выделением факторов: ГЕНОТИП, ПОЛ, ВОЗРАСТ и ЛАТЕРАЛЬНОСТЬ (предъявление в правое ухо, в левое ухо). Выявлена значимость всех выделенных факторов и взаимодействия ВОЗРАСТ х ЛАТЕРАЛЬНОСТЬ (F(1, 313) = 8.178, p = 0.004). Значимость фактора ГЕНОТИП (F(2, 313) = 3.170, p = 0.043) обусловлена более эффективным воспроизведением слов носителями генотипа 12/12 по сравнению с гетерозиготным генотипом (рис. 1 (а)), ПОЛ (F(1, 313) = 8.78, p = 0.003) – женщинами по сравнению с мужчинами, ВОЗРАСТ (F(1, 313) = 94.747, p = 0.000) – молодыми по сравнению с пожилыми, ЛАТЕРАЛЬНОСТЬ (F(1, 313) = 183.770, p = 0.000) –лучшим воспроизведением слов, адресованных речевому левому полушарию. В основе взаимодействия ВОЗРАСТ × ЛАТЕРАЛЬНОСТЬ лежит более выраженное возрастное снижение воспроизведения слов с левого по сравнению с правым ухом.
Рис. 1.
Эффективность воспроизведения слов в дихотическом тесте (а) и узнавания предъявленных для запоминания фигур (б, в) в зависимости от генотипов полиморфизма STin2VNTR гена транспортера серотонина. Вертикальные отрезки – ошибки среднего, на каждом рисунке одинаковыми значками обозначены достоверно отличающиеся значения, на рис. (в) сплошная линия – молодые, пунктирная – пожилые испытуемые.
Fig. 1. Efficiency of retrieval of words in the dichotic test (а) and recognition of figures presented for memorization (б, в) depending on the genotypes of the STin2VNTR polymorphism of the serotonin transporter gene. Vertical lines are standard errors of the mean, in each figure the same marks indicate significantly different values, in Fig. (в) the solid line is the young, the dotted line is the elderly subjects.
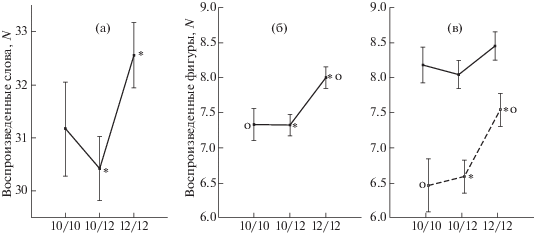
Дисперсионный анализ узнавания предъявленных для запоминания слогов и фигур был проведен для факторов ГЕНОТИП, ПОЛ, ВОЗРАСТ и ТИП ЗАДАНИЯ (слоги, фигуры). Анализ выявил значимость фактора ВОЗРАСТ ( F(1, 321) = 51.579, p = 0.000) и взаимодействий ПОЛ × ТИП ЗАДАНИЯ (F(1, 321) = 4.317, p = 0.038), ГЕНОТИП × ТИП ЗАДАНИЯ (F(2, 321) = 3.435, p = 0.033) и ГЕНОТИП × ВОЗРАСТ × ТИП ЗАДАНИЯ (F(2, 321) = 4.528, p = 0.012). Взаимодействие первых двух факторов обусловлено лучшим запоминанием фигур по сравнению со слогами мужчинами при отсутствии таких различий у женщин. Поскольку, кроме фактора ПОЛ, все остальные факторы вошли в значимое взаимодействие, целесообразно рассмотреть их вклад в это взаимодействие. В задании с запоминанием слогов не обнаружено связанных с генотипом STin2VNTR различий в эффективности запоминания.
При анализе запоминания фигур выявлена значимость фактора ГЕНОТИП (F(1, 321) = = 10.670, p = 0.001), обусловленная меньшим числом ошибок у носителей генотипа 12/12 по сравнению с носители аллеля 10' (рис. 1 (б)). При одинаковой направленности влияния генотипов Stin2VNTR на эффективность памяти у молодых и пожилых испытуемых, только в старшей возрастной группе существовали достоверные различия в эффективности памяти между 12/12 и носителями аллеля 10' (с генотипом 10'/10', р = 0.016 и 10'/12, р = 0.005; (рис. 1 (в)).
ОБСУЖДЕНИЕ РЕЗУЛЬТАТОВ
Частота встречаемости аллеля 12 в нашей выборке (0.60) соответствует результатам других исследований, оценивающих встречаемость аллеля 12 у европейцев в пределах 0.54–0.66 [Fan, Sklar, 2005].
Проведенное исследование выявило ассоциации полиморфизма STin2VNTR гена транспортера серотонина с эффективностью вербальной памяти, определяемой по воспроизведению существительных в процедуре дихотического тестирования. Наилучшие показатели запоминания продемонстрировали носители гомозиготного генотипа 12/12. Нам удалось найти лишь несколько работ, в которых рассматриваются ассоциации между рассматриваемым полиморфизмом и памятью. Только в одной из них были исследованы эффекты полиморфизма на показатели памяти у здоровых испытуемых. В исследовании участвовали только пожилые люди без когнитивных нарушений, и рассматривалось влияние STin2VNTR как на исходные показатели когнитивных функций, так и динамику их возрастного снижения [Payton et al., 2005]. В отличие от полученных нами результатов авторы не обнаружили влияния STin2VNTR на показатели когнитивных функций пожилых людей при первом тестировании, однако выявили более быстрое их снижение у носителей генотипа 12/12 по сравнению с 10/10 и гетерозиготным генотипом в течение последующих 15 лет после начала исследования. Различия были достоверны для показателей флюидного интеллекта, семантической памяти и лишь близки к достоверности при непосредственном и отсроченном воспроизведении вербальной информации. Приведенные данные указывают на особенности влияния STin2VNTR в зависимости от тестируемых показателей памяти. В этой связи следует отметить, что применявшаяся в нашем исследовании процедура дихотического исследования значительно отличается от проводимых в цитируемой работе тестов. Кроме этого, во многих исследованиях показано значение внешних факторов в определении характера взаимодействия функциональных полиморфизмов с поведенческими характеристиками. В нашем исследовании все испытуемые составляли группу социально активных и ментально здоровых людей. Остальные обнаруженные нами работы проведены без дифференциации возраста испытуемых и выполнены на больных с аффективными расстройствами. Отмечено, что носители генотипа 12/12 обладают более высокими показателями памяти по сравнению с носителями аллеля 10 полиморфизма STin2VNTR [Dömötör et al., 2007; Sarosi et al., 2008]. Эти результаты согласуются с нашими данными, которые независимо от возраста выявили общий эффект: лучшую память у носителей генотипа 12/12 по сравнению с другими генотипами. Однако встает вопрос о сопоставимости результатов, полученных на здоровых испытуемых и лицах с аффективной патологией, что требует дополнительных исследований.
Что касается влияния STin2VNTR на показатели воспроизведения предъявленных для запоминания слогов и фигур, то генетические различия выявлены только при запоминании фигур, что подтверждают данные нашего предыдущего исследования, направленного на выявление эффектов другого функционального полиморфизм гена транспортера серотонина – 5-HTTLPR [Вольф и др., 2017]. Однако сравнение эффектов двух полиморфизмов показывает, что для полиморфизма 5-HTTLPR наилучшие показатели воспроизведения обнаружены у носителей S/S генотипа, характеризующегося низкой экспрессией гена транспортера серотонина, а для STin2VNTR – у носителей генотипа 12/12, считающегося наиболее сильным усилителем экспрессии. Противоречивость полученных результатов может быть обусловлена тем, что изучение экспрессии полиморфизмов гена транспортера серотонина проводилось в основном in vitro. Возможность различной регуляции экспрессии генов в мозге и на периферии показана во многих исследованиях. Для менее изученного полиморфизма STin2VNTR единственное исследование его влияния на доступность транспортера серотонина в мозге показало связь генотипа 12/12 не с усилением, как это установлено in vitro [Hranilovic et al., 2004], а с ослаблением доступности транспортера [Bah et al., 2008]. Наряду с этим также обнаружено, что аллель 12 полиморфизма STin2VNTR, как и S аллель полиморфизма 5-HTTLPR, ассоциирован с личностной тревожностью, что позволяет предполагать сходное влияние этих полиморфизмов на серотонинергическую систему мозга [Melke et al., 2001]. Таким образом, полученные нами результаты согласуются с данными о сходстве эффектов 12 и S аллелей, то есть их возможной связи с пониженной экспрессией транспортера. При наличии общего эффекта генотипа, указывающего на однонаправленность ассоциаций эффективности запоминания с полиморфизмом STin2VNTR у испытуемых разного возраста, достоверные генетические различия выявлены только у пожилых людей. При исследовании обусловленных старением изменений доступности транспортера серотонина в мозге показано ее возрастное снижение [Kakiuchi et al., T, 2001]. Возможно, более низкий при старении уровень доступности гена транспортера серотонина способствует контрастированию влияния генотипов, в разной степени усиливающих его экспрессию, и связанных с ними поведенческих эндофенотипов.
Что касается выявленных в работе особенностей памяти, связанных с полом и возрастом, то они согласуются с широко известными данными о лучших вербальных способностях у женщин и образных у мужчин [Andreano, Cahill, 2009] и о снижении памяти при старении [Park, Festini, 2017].
Полученные в работе результаты имеют ряд ограничений, требующих обсуждения. Во-первых, наблюдаемые ассоциации с показателями памяти могут зависеть не только от изучаемого полиморфизма STin2VNTR, но накладываться на эффекты, обусловленные другим функциональным полиморфизмом 5-HTTLPR, гена транспортера серотонина. В частности, эффекты полиморфизма могут зависеть от неравновесного сцепления между полиморфными системами STin2VNTR и 5-HTTLPR. Кроме этого, эффект полиморфизма может зависеть от половых различий и эпигенетического влияния факторов окружающей среды, то есть контекста реализации генотипа. Такое влияние на ген транспортера серотонина отмечено во многих исследованиях [Belsky, Pluess, 2009]. В то же время следует отметить некоторые сильные стороны нашего исследования. Во-первых, оно выполнено на относительно большой выборке. Во-вторых, в исследовании проведено сравнение эффектов в зависимости от пола и возраста испытуемых, факторов, влияющих на фенотипические проявления генотипа. И наконец, одним из методологических преимуществ нашего исследования является социальная однородность выборки, в которой представлена популяция ментально здоровых и успешных молодых и пожилых людей, относящихся к кавказской группе народов. Особо следует отметить, что все испытуемые старшего возраста продолжали профессиональную деятельность на момент исследования, что является важнейшим показателем “успешного” ментального старения. Такая однородность популяции при сравнении с накапливающимися данными других исследований может иметь существенное значение для дифференцировки влияния генетических и эпигенетических факторов в эффектах полиморфных генотипов гена транспортера серотонина в отношении когнитивных функций.
ВЫВОДЫ
1. Полиморфизм STin2VNTR гена транспортера серотонина ассоциирован с эффективностью запоминания слов в дихотическом тесте: носители генотипа 12/12 запоминали больше слов по сравнению с носителями аллелeй 10'.
2. При узнавании предъявлявшихся для запоминания слов и фигур полиморфизм STin2VNTR был ассоциирован только с запоминанием фигур.
3. Носители генотипа 12/12 воспроизводили большее число фигур по сравнению с носителями аллелей 10', эффект генотипа был более выражен у пожилых испытуемых.
Исследование выполнено при финансовой поддержке РФФИ и Правительства Новосибирской области в рамках научного проекта №17-46-540705.
Список литературы
Вольф Н.В., Базовкина Д.В., Куликов А.В. 5-HTTLPR полиморфизм гена транспортера серотонина и интеллектуальная среда профессиональной деятельности как факторы, ассоциированные с изменениями памяти при старении. Журн. высш. нерв. деят. 2017. 67 (6): 744–754.
Пучкова А.Н., Дорохов В.Б. Молекулярно-генетические исследования индивидуальных отличий и профессиональная деятельность Журн. высш. нервн. деят. 2015. 65 (2): 188–202.
Andreano J.M., Cahill L. Sex influences on the neurobiology of learning and memory. Learn Mem. 2009.16 (4): 248–266.
Avula R., Rand A., Black J.L., O’Kane D.J. Simultaneous genotyping of multiple polymorphisms in human serotonin transporter gene and detection of novel allelic variants. Transl. Psychiatry. 2011.1:e32.
Bah J., Lindstrom M., Westberg L., Manneras L., Ryding E., Henningsson S., Melke J., Rosén I., Träskman-Bendz L., Eriksson E. Serotonin transporter gene polymorphisms: effect on serotonin transporter availability in the brain of suicide attempters. Psychiatry Res. 2008. 162 (3): 221–229.
Belsky J., Pluess M. Beyond diathesis stress: Differential susceptibility to environmental influences. Psychol. Bull. 2009. 135: 885–908.
Borg J., Henningsson S., Saijo T., Makoto I., Bah J., WestbergL., et al. Serotonin transporter genotype is associated with cognitive performance but not regional 5-HT1A receptor binding in humans. Int. J. Neuropsychopharmacol. 2009. 12 (6): 783–792.
Dömötör E., Sárosi A., Balogh G., Székely A., Héjjas K., Sasvári-Székely M., Faludi G. Association of neurocognitive endophenotype and STin2 polymorphism in major depressive disorder. Neuropsychopharmacol. Hung. 2007. (2): 53–62.
Fan J.B., Sklar P. Meta-analysis reveals association between serotonin transporter gene STin2 VNTR polymorphism and schizophrenia. Mol. Psychiatry. 2005. 10 (10): 928–938.
Fiskerstrand C.E., Lovejoy E.A., Quinn J.P. An intronic polymorphic domain often associated with susceptibility to affective disorders has allele dependent differential enhancer activity in embryonic stem cells. FEBS Lett. 1999. 458 (2): 171–174.
Homberg J.R., Molteni R., Calabrese F., Riva M.A. The serotonin BDNF duo: developmental implications for the vulnerability to psychopathology. Neurosci. Biobehav. Rev. 2014. 43: 35–47.
Hranilovic D., Stefulj J., Schwab S., Borrmann-Hassenbach M., Albus M., Jernej B., Wildenauer D. Serotonin transporter promoter and intron 2 polymorphisms: relationship between allelic variants and gene expression. Biol Psychiatry. 2004. 55 (11): 1090–1094.
Kakiuchi T., Tsukada H., Fukumoto D., Nishiyama S. Effects of aging on serotonin transporter availaility and its response to fluvoxamine in the living brain: PET study with [ (11)C] (+)McN5652 and [ (11)C] (-)McN5652 in conscious monkeys. Synapse. 2001. 40 (3): 170–179.
Madsen K., Errizoe D., Mortensen E.L., Gade A., Madsen J., Baaré W., Knudsen G.M., Hasselbalch S.G. Cognitive function is related to frontostriatal serotonin transporter levels – A brain PET study in young healthy subjects. Psychopharmacology. 2011. 213 (2-3): 573–581.
Melke J., Landén M., Baghei F., Rosmond R., Holm G., Björntorp P., Westberg L., Hellstrand M., Eriksson E. Serotonin transporter gene polymorphisms are associated with anxiety-related personality traits in women. Am J Med Genet. 2001. 105 (5): 458–463.
Meneses A. 5-HT system and cognition. Neurosci. Biobehav. Rev. 1999. 23 (8): 1111–1125.
Miller S.A., Dykes D.D., Polesky H.F. A simple salting out procedure for extracting DNA from human nucleated cells. Nucleic Acids Res. 1988. 16 (3): 1215.
Park D.C., Festini S.B. Theories of Memory and Aging: A Look at the Past and a Glimpse of the Future. J. Gerontol B. Psychol. Sci. Soc. Sci. 2017. 72 (1): 82–90.
Payton A., Gibbons L., Davidson Y., Ollier W., Rabbitt P., Worthington J., Pickles A., Pendleton N., Horan M. Influence of serotonin transporter gene polymorphisms on cognitive decline and cognitive abilities in a nondemented elderly population. Mol. Psychiatry. 2005. 10 (12): 1133–1139.
Roiser J., Muller U., Clark L., Sahakian B.J. The effects of acute tryptophan depletion and serotonin transporter polymorphism on emotional processing in memory and attention. Int. J. Neuropsychopharmacol. 2007. 10 (4): 449–461.
Sarosi A., Gonda X., Balogh G., Domotor E., Szekely A., Hejjas K., Sasvari-Szekely M., Faludi G. Association of the STin2 polymorphism of the serotonin transporter gene with a neurocognitive endophenotype in major depressive disorder. Prog. Neuropsychopharmaco.l Biol. Psychiatry. 2008. 32 (7): 1667–1672.
Sodhi M.S., Sanders-Bush E. Serotonin and brain development. Int. Rev. Neurobiol. 2004. 59: 111–174.
van Kesteren R.E., Spencer G.E. The role of neurotransmitters in neurite outgrowth and synapse formation. Rev Neurosci. 2003. 14 (3): 217–231.
Дополнительные материалы отсутствуют.
Инструменты
Журнал высшей нервной деятельности им. И.П. Павлова