

Журнал высшей нервной деятельности им. И.П. Павлова, 2019, T. 69, № 5, стр. 590-600
ПОВЕДЕНИЕ ДОМЕСТИЦИРУЕМЫХ И АГРЕССИВНЫХ ЛИСИЦ В СИТУАЦИИ ВЫБОРА МЕЖДУ РАЗНЫМИ КОЛИЧЕСТВАМИ КУСОЧКОВ ПИЩИ
И. А. Мухамедшина 1, *, А. В. Харламова 1, **, Л. Н. Трут 1
1 Федеральное государственное бюджетное научное учреждение “Федеральный исследовательский центр
Институт цитологии и генетики Сибирского отделения Российской академии наук” (ИЦиГ СО РАН)
Новосибирск, Россия
* E-mail: aden_66@mail.ru
** E-mail: kharlam@bionet.nsc.ru
Поступила в редакцию 27.12.2017
После доработки 24.10.2018
Принята к публикации 20.11.2018
Аннотация
В настоящей работе оценивали поведение лисиц, прошедших длительный отбор на доместикационное и агрессивное поведение по отношению к человеку, а также у лисиц, не подвергавшихся специальному отбору по поведению в тестах на выбор между двумя емкостями, содержащими различное количество одинаковых по размеру кусочков пищи. Лисицам предъявляли следующие соотношения между количествами кусочков пищи: 10 и 1, 5 и 1, 10 и 5, 3 и 2. Большее количество предъявляли с одной и той же стороны до тех пор, пока животное не сделает пять правильных выборов (то есть, выборов большего количества) подряд. Поэтому задача выглядела как последовательная переделка навыка выбирать корм то с одной, то с другой стороны. Когда различия между количествами кусочков было максимальными (10 и 1), 46% ручных, 38% неселекционированных и 36% агрессивных лисиц достоверно неслучайно выбирали большее количество кусочков пищи (различия между популяциями недостоверны). В выборе между 5 и 1 кусочками доля ручных лисиц, неслучайно выбиравших большее количество, оказалась достоверно выше по сравнению с агрессивными и неселекционируемыми. При повышении сложности задачи (выбор между 5 и 1 кусочками) агрессивные и неселекционируемые лисицы чаще, чем ручные, начинают выбирать емкость на какой-либо одной из сторон (либо только правую, либо левую), независимо от числа кусочков в ней. При дальнейшем усложнении задачи (соотношение между количествами кусочков 10 и 5, либо 3 и 2) у лисиц из всех групп возрастает процент односторонних выборов. В статье описаны и обсуждаются особенности поведения животных, которые могли явиться причиной наблюдаемых различий.
ВВЕДЕНИЕ
Изучение различных параметров когнитивных способностей животных является актуальной научной задачей и широко изучается большим кругом исследователей [Banerjee et al., 2009; MacLean et al., 2014; Broadway et al., 2017]. По мнению К. Лоренца, развитие когнитивных способностей является необходимым условием в процессе приспособления животного к изменениям окружающей среды [Лоренц, 1998]. Выбор стратегии поведения следует признать важным принципом адаптации к окружающей среде и к выживанию, что делает этот принцип актуальным для исследования когнитивного поведения. Поэтому исследователи поведения стремятся к поиску таких методик, которые предоставляли бы животному возможность самостоятельно выбирать стратегию при решении той или иной задачи. К ним относится, например, изучение поведения животных в лабиринтах, традиционно используемые исследователями когнитивного поведения [Tryon, 1940; Lipp et al., 2001; Плескачева, 2008] и исследование способности животных к различению количественных соотношений [Зорина, Смирнова, 1994, 1996; Резникова, 2004; Reznikova, Ryabko, 2011].
В исследованиях способностей животных к пространственному обучению (A-not-B detour task) на собаках, лошадях, ослах и мулах было показано, что животные в ситуации свободного выбора направления движения стремятся раз за разом обходить барьер с той стороны, которая подкреплялась в первом тесте, даже несмотря на последующее отсутствие подкрепления [Osthaus et al., 2010, 2013]. Такую форму поведения авторы назвали устойчивой пространственной ошибкой (spatial perseveration error), и у некоторых собак настойчивые попытки обойти ширму с одной и той же стороны возникали уже после одного единственного подкрепления предпочитаемой стороны, даже в тех вариантах эксперимента, когда животное через прозрачный барьер могло видеть подкрепление, размещенное с другой стороны [Osthaus et al., 2010]. Авторы объясняют такое поведение собак очень быстрым и прочным формированием у них условно-рефлекторных связей и возможным эффектом доместикации. В сравнительном исследовании собак и волков в задаче A-not-B [Topál et al., 2009] волки были менее склонны к настойчивым повторениям ошибочных выборов. Авторы сравнительного исследования объясняют большую склонность собак к односторонним выборам в задаче A-not-B их большей чувствительностью к коммуникативным сигналам человека, приобретенной в процессе их одомашнивания [Topál et al., 2009]. Согласно их гипотезе, собаки совершают повторяющуюся ошибку (perseverative bias) вследствие неправильной интерпретации коммуникативных сигналов, подаваемых экспериментатором.
Существуют также и другие интерпретации причин односторонних выборов, совершаемых различными животными в когнитивных тестах. Например, односторонние выборы в A-not-B task и других подобных тестах считаются показателем слабого самоконтроля (так называемый inhibitory control, который проявляется в низкой способности животных затормозить форму поведения, не являющуюся адаптивной) [Brucks et al., 2017].
В другом исследовании авторы обучали крыс выбирать направление движения в рукавах радиального лабиринта [Lee, Solivan, 2008; 2010; Lee, Kim, 2010; Jo, Lee, 2010]. Каждый из рукавов заканчивался двумя выходами. В зависимости от номера рукава крыса должна была либо повернуть в направлении подкрепляемого объекта (object in place strategy), либо в направлении подкрепляемой стороны рукава, независимо от находящегося там объекта (location-in-place strategy). Исследователи пишут, что прежде чем обучиться выбирать направление движения, ориентируясь на находящийся в рукаве объект, крысы сначала демонстрировали односторонние выборы [Lee, Kim, 2010]. В этом исследовании односторонние выборы рассматриваются авторами как стратегия поведения.
Стремление животных выбирать емкость с кормом с одной и той же стороны наблюдается и в решении животными задач на различение количественных факторов, в таких исследованиях эту форму поведения называют “латеральными предпочтениями” (laterality bias) [Petrazzini, Wynne, 2016; 2017, Macpherson et al., 2013, Range et al., 2014; Banszegi et al., 2016]. Латеральные предпочтения наблюдаются у разных животных на разных этапах эксперимента – как в предварительном тесте, где в одной емкости предъявляется один кусочек пищи, а другая емкость остается пустой (1 vs 0 condition), так и в основном тесте, когда две емкости содержат разное количество кусочков. И хотя в тесте A-not-B волки были не склонны совершать односторонние выборы [Topál et al. 2009], в тесте на способность к различению количественных соотношений некоторые волки демонстрировали настойчивые односторонние выборы даже в самом простом варианте предлагаемой задачи [Petrazzini, Wynne, 2017]. Односторонние выборы наблюдались и при исследовании способности серых ворон к относительным количественным оценкам, и авторы объясняли это поведение проявлением невротического состояния у птиц при увеличении сложности задачи [Зорина, Смирнова, 1996].
В данном исследовании мы ставим цель исследовать особенности поведения лисиц при решении ими задач на различение количественных соотношений. Ранее нами было начато исследование когнитивных способностей серебристо-черных лисиц (Vulpes vulpes) из экспериментальных популяций, подвергавшихся длительному отбору по поведению в двух контрастных направлениях. Ручные в течение более 50 поколений отбирались на дружелюбное отношение к человеку. Агрессивные лисицы прошли не менее 40 поколений отбора на усиление агрессивного реагирования на человека [Trut et al., 2009].
Нами было показано, что отбор по реакциям на человека затронул некоторые параметры когнитивного поведения и высшей нервной деятельности лисиц – агрессивные способны дольше фокусировать взгляд на объекте пищевого подкрепления, а у ручных слабее развита способность к угашению выработанного поведения [Мухамедшина и др., 2014], выше уровень общей возбудимости и, предположительно, выше подвижность нервных процессов и пластичность поведения [Мухамедшина и др., 2019]. Показано также, что у ручных лисиц по сравнению с неселекционируемыми по поведению выше скорость обучения в челночной камере и понижен порог нервно-мышечной возбудимости [Васильева, 1991а, b].
Настоящая работа является продолжением исследования влияния отбора по поведению на некоторые когнитивные способности лисиц. Главная цель данной работы состоит в том, чтобы оценить поведение лисиц при предъявлении им задачи по дифференцированию количеств и сравнить особенности поведения ручных, агрессивных и неселекционированных лисиц в этом эксперименте.
МЕТОДИКА
Место проведения работы и животные – объект исследования
Работа проводилась на экспериментальной звероферме ИЦиГ СО РАН (г. Новосибирск), в качестве объекта использовали животных из популяций, селекционируемых на ручное и агрессивное поведение по отношению к человеку. Все животные, включенные в эксперимент, были предварительно протестированы по стандартной методике оценки как ручного, так и агрессивного поведения [Трут, 1980; Trut, 1999], в эксперимент были взяты наиболее контрастные животные, с максимальным проявлением агрессии или дружелюбия. В исследование также была включена выборка контрольных лисиц, содержащихся в тех же условиях, имеющих такой же опыт контактов с человеком на протяжении всей жизни, но не подвергавшихся систематическому отбору по поведению.
Работа проводилась в течение двух последовательных лет (2015–2016 гг.). В 2015 г. в исследование были включены животные в возрасте полутора лет, а в 2016 г. – лисицы в возрасте 6–7 мес., в равном соотношении самцы и самки. Общее количество предварительно отобранных животных: 47 ручных, 39 агрессивных и 36 неселекционируемых. В ходе тестирования не все они оказались способны выполнить условия теста, некоторые были исключены на разных этапах эксперимента (подробнее см. в разделе Результаты), в итоге были получены данные для 28 ручных, 33 агрессивных и 31 неселекционируемых лисиц. Лисицы содержались в стандартных условиях, обычных для коммерческих звероферм, в клетках размером 90 × 90 × 90 см, расположенных в два ряда под общим навесом, по 80 клеток в ряду. Крайне важно подчеркнуть, что данная работа проводилась в условиях производственного содержания, в домашней клетке, поскольку любое перемещение из привычных условий, или помещение перед домашней клеткой объемной ширмы, скрывающей экспериментатора, повлияло бы на поведение лисиц сильнее, чем присутствие человека, к которому все лисицы привыкли в одинаковой степени. Поскольку основной идеей эксперимента было изменение поведения путем отбора [Беляев, 1983], контакты с человеком для ручных, агрессивных и неселекционируемых по поведению лисиц ограничены процедурами ухода и кормления. Емкости с кормом вдвигали в ту же прорезь в клетке, в которой обычно располагается “домашняя” кормушка. Корм работница выдает рукой, т.е. все манипуляции в ходе теста были максимально приближены к той ситуации, которая ежедневно происходит на ферме при кормлении животных.
Перед началом эксперимента все отобранные для тестирования животные были рассажены так, что соседние клетки в ряду, справа и слева от каждой тестируемой лисицы, оставались пустыми, чтобы исключить отвлекающее влияние лисиц, сидящих в соседних клетках и возбуждающихся при виде подкормки, на поведение животного во время теста. На адаптацию к смене домашней клетки животным давали две недели, после этого проводили тестирование.
Все тесты проводили в августе–октябре. Поскольку в данном эксперименте лисицам предлагалось выбрать между разными количествами кусочков пищи, необходимым условием была пищевая мотивация животных к получению подкормки, поэтому тест проводили после суточной пищевой депривации. После прохождения теста все животные получали суточную порцию корма, независимо от успешности решения задачи.
Процедура тестирования
Лисице через прорезь в домашней клетке одновременно (на виду у животного) вдвигали две одинаковые емкости, содержащие разное количество одинаковых по размеру кусочков вареной печени, и давали возможность съесть кусочки печени только из той емкости, которая была ею выбрана.
Как только лисица выбирала одну из емкостей с кормом, экспериментатор сразу же убирал вторую. Всего каждому животному предъявляли четыре задачи, в порядке усложнения: 10:1, 5:1, 10:5 и 3:2. Известно, что способность к различению количественных соотношений подчиняется закону Вебера-Фехнера [Gallistel, Gelman, 2000; Lewis et al., 2005]. Согласно этому закону сложность задачи на распознавание повышается при уменьшении соотношения и при увеличении значений сравниваемых количеств. Поэтому мы исходили из предположения, что наиболее простыми для животных будут те задачи, в которых в одной из мисок находился только один кусочек печени, т.е. когда лисице нужно было сделать выбор между единственным кусочком и несколькими, а более сложными – те, в которых предъявляли либо большое количество кусочков как справа, так и слева (10 и 5), либо разница между двумя предъявляемыми количествами была минимальной (3 и 2). В один день животному предъявляли только одну задачу, в течение теста ее предъявляли 30 раз. Интервал между тестами составлял семь дней, по истечении которых животному предъявляли следующую по сложности задачу. Поскольку голодное животное мотивировано на возможность получить больше корма, выбор емкости с большим количеством кусочков пищи мы принимали за правильный. Общепринято, что задачи на различение количеств строятся с предъявлением большего количества с левой и правой сторон в квази-случайном порядке [Ward, Smuts, 2007; Hunt et al., 2008; Hanus, Call, 2007]. Ранее мы проводили с лисицами тест по этой схеме (неопубликованные данные), и при решении задачи выбора большего количества мы наблюдали поведение лисиц, которые предпочитали раз за разом выбирать емкость с кормом на одной стороне (либо справа, либо слева), независимо от количества корма в ней. В контексте данного исследования для выявления особенностей поведения лисиц при выборе разного количества корма такие животные представляют отдельный интерес. Поэтому мы поставили получение ими большего количества в зависимость от их предыдущего выбора. Мы модифицировали тест, предъявляя большее количество кусочков корма на одной и той же стороне до достижения критерия пяти “правильных” выборов подряд. Только после этого большее количество начинали предъявлять на другой стороне и делали это также до достижения пяти “правильных” выборов подряд. Таким образом, фактически наша задача была сведена к переделке навыка выбирать корм с одной из сторон на выбор корма с другой стороны. (видеозапись доступна на сайте: http://www.bionet.nsc.ru/razrabotki/prikladnyie-razrabotki/zhivotnovodstvo/lisiczyi/ruchnaya-lisicza-vyibiraet-bolshee-v-sootnoshenii-10-k-1.html). Если лисица начинала предпочитать одну сторону и выбирала при этом меньшее количество, то она неизбежно получала меньшее количество до тех пор, пока не сменяла сторону выбора. Такую форму поведения мы назвали “латеральные предпочтения” (видеозапись доступна на сайте: http://www.bionet.nsc.ru/razrabotki/prikladnyie-razrabotki/zhivotnovodstvo/lisiczyi/agressivnaya-lisicza-vyibiraet-emkost-s-odnoj-storonyi-nezavisimo-ot-kolichestva-kusochkov-pishhi.html).
Кроме того, для выявления возможных предпочтений отдельными лисицами правой или левой стороны мы провели два дополнительных теста с теми же лисицами.
В первом из них экспериментатор на виду у лисицы последовательно помещал в две миски одинаковое количество кусочков лакомства (4:4), затем миски, как и в основном эксперименте, вдвигались в прорезь домашней клетки животного, и лисице предоставлялась возможность сделать выбор (и съесть корм, находящийся в выбранной емкости). Как только лисица делала выбор, вторую емкость с кормом сразу же выдвигали из клетки. Так как животному предъявляли одинаковое количество кусочков на каждой из сторон, любой выбор можно было оценить как правильный. В данном тесте мы ставили цель оценить латеральные предпочтения лисиц. Поэтому он был ограничен по времени (с каждым животным он проводился в течение 7 мин), а не по числу предъявлений. За это время экспериментатор предъявлял лисице задачу около 20 раз.
Через день с теми же лисицами проводили тест с пустой миской. Экспериментатор помещал в одну миску 4 кусочка вареной печени, другая же миска оставалась пустой, но при этом экспериментатор делал четыре обманных движения, имитируя выкладывание четырех кусочков во вторую миску (таким образом, количество кусочков в мисках составляло 4:0). Если лисица выбирала пустую миску, то в следующем предъявлении эта миска вновь оставалась пустой, по аналогии со схемой предъявления в основном тесте, когда экспериментатор повторял одно и то же предъявление до тех пор, пока лисица не выбирала емкость с большим количеством кусочков. Продолжительность данного теста, как и предыдущего, составляла 7 мин. Все тесты видеорегистрировались, параметры поведения извлекались экспериментатором из видеозаписи.
Статистическая обработка была осуществлена в пакете программ Statistica 6. Для сравнения групп по исследованным показателям поведения (процент выборов большего количества; процент выборов пустой миски; процент “латерализации”; число смен стороны предъявления большего количества; доля лисиц, достоверно выбирающих большее количество; доля лисиц, исключенных в первый день тестирования) использовали непараметрические критерии Манна–Уитни (Mann_Whitney U test) и 2 × 2 Фишера (F-критерий). Достоверность отличия от случайного выбора 50:50 оценивали с использованием критерия falsh discovery rate (FDR) (Benjamini, Hochberg, 1995). Влияние года (т.е. правомерность отнесения животных, протестированных в разные годы, к одной выборке) проверялось с помощью однофакторного дисперсионного анализа (ANOVA one-way). Различия считали достоверными при p ≤ 0.05.
Вся работа с животными проводится в соответствии с международными нормами охраны животных, используемых в научных целях (Директива 2010/63/EU Европейского Парламента и Совета Европейского Союза).
РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЯ
Полученные в работе результаты показывают, что лисицы демонстрируют достаточно большую вариабельность в решении предъявленных тестов, которая оказалась во многом связанной с принадлежностью лисицы к доместицированной, агрессивной или “неселекционированной” группе.
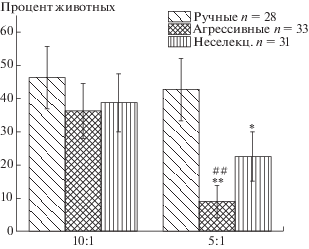
Сравнение результатов выполнения задач лисицами из трех групп показало следующее. В решении самой простой задачи, когда различия между предъявляемыми количествами кусочков корма максимальное (10 и 1), значительная часть протестированных лисиц оказались способны уловить закономерность предъявления задачи и превзошли критерий FDR не-случайности выбора большего количества. Не обнаружено достоверных различий между группами по доле животных, успешно решивших задачу 10:1 (рис. 1). В задаче 5:1 доля ручных лисиц, неслучайно выбиравших большее количество, оказалась достоверно выше по сравнению с агрессивными и неселекционируемыми (рис. 1). С усложнением задачи снижается доля животных, успешно выбирающих большее количество. В соотношениях 10:5 и 3:2 ни одна протестированная лисица из всех трех групп не превзошла строгий критерий неслучайности выбора большего количества FDR.
Рис. 1.
Процент лисиц, которые достоверно неслучайно выбирали большее количество кусочков печени в соотношениях 10:1 и 5:1. # – p < < 0.01 по сравнению с 10:1. * – p < 0.05; ** – p < < 0.01 по сравнению с ручными лисицами.
Fig. 1. The percent of foxes, who choose more quantity of liver pieces significantly above chance in tasks 10:1 and 5:1. # – p < 0.01 in comparison with 10:1. * – p < 0.05; ** – p < 0.01 in comparison with tame foxes.
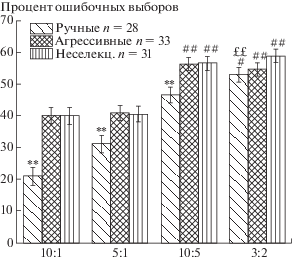
Определенный нами порядок предъявления единиц подкрепления приводил к тому, что когда лисица раз за разом выбирала одну сторону, она могла соответственно выбрать меньшее количество кусочков печени несколько раз подряд. Доля таких повторяющихся односторонних выборов меньшего количества (т.е. суммарное количество выборов во время “латеральных предпочтений”, отнесенное к общему числу предъявлений задачи) была различной в группах лисиц и в задачах разной сложности (рис. 2), из которого видно, что она увеличивается с усложнением задачи. Однако в целом ручные лисицы достоверно реже (рис. 2) выбирают меньшее количество кусочков печени несколько (более двух) раз подряд – по сравнению с неселекционируемыми на протяжении всех тестов (10:1, 5:1, 10:5, 3:2) и по сравнению с агрессивными – на протяжении всех тестов, кроме 3:2.
Рис. 2.
Процент ошибочных выборов во время латеральных предпочтений в соотношениях 10:1, 5:1, 10:5 и 3:2. # – p < 0.05; ## – p < 0.01 по сравнению со случайным выбором 50:50. ** – p < 0.01 по сравнению с агрессивными и неселекционируемыми лисицами. ££ p ≤ 0.01 по сравнению с неселекционируемыми лисицами.
Fig. 2. Percent of wrong choices during uni-lateral choices in tasks 10:1, 5:1, 10:5 and 3:2. # – p < 0.05; ## – p <0.01 in comparison with choice “by chance” – 50:50. ** – p < 0.01 in comparison with aggressive and unselected foxes. ££ p ≤ 0.01 in comparison with unselected foxes.
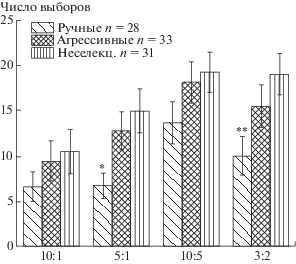
На рисунке 3 отражен процент выборов во время латеральных предпочтений для самой простой задачи (10:1) отдельно у лисиц, которые достоверно неслучайно выбирали большее количество кусочков, и у всех остальных протестированных животных для каждой группы. На этой гистограмме видно, что показатель латерализации у успешных лисиц не различается между тремя исследованными выборками и в среднем не превышает 15%. Это значение достоверно меньше, чем у остальных (неуспешных) протестированных животных, для которых показаны различия между группами по этому показателю. Средний процент латерализации у ручных лисиц, которые не достигли критерия неслучайного выбора, оказался достоверно ниже, чем у агрессивных и неселекционируемых лисиц. Такая особенность поведения ручных лисиц приводит к тому, что они достигают критерия пяти правильных выборов подряд быстрее, чем неселекционируемые лисицы (достоверно) и агрессивные (на уровне тенденции) (рис. 4).
Рис. 3.
Процент ошибочных выборов во время латеральных предпочтений в тесте с соотношением кусочков 10:1 для лисиц, которые достоверно неслучайно выбирали большее количество (успешные, У) в сравнении с лисицами, у которых выбор большего достоверно не отличался от случайного (неуспешные, Н). *** p ≤ ≤ 0.001 по сравнению с агрессивными и неселекционируемыми лисицами. ### – p < 0.001 разница между “успешными” и “неуспешными”.
Fig. 3. Percent of wrong choices during uni-lateral choices in task 10:1 – separately for foxes who choose more quantity significantly above chance (“successful”) and “unsuccessful” (who choose quantity “by chance”). *** p ≤ 0.001 in comparison with aggressive and unselected foxes. ### – p < < 0.001 in comparison with “unsuccessful”.
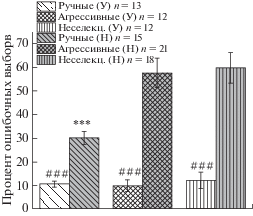
Рис. 4.
Число предъявлений задачи на одной и той же стороне до первых пяти правильных выборов подряд.* – p <0.05; ** – p <0.01 по сравнению с неселекционируемыми лисицами.
Fig. 4. The number of task submissions on the same side before firstly reaching of five correct choices. * – p < 0.05; ** – p < 0.01 in comparison with unselected foxes.
В эксперименте, когда лисицам предлагалось выбрать между пустой миской и четырьмя кусочками печени (4:0), процент выборов пустой миски оказался достоверно ниже у неселекционируемых по поведению лисиц, чем у ручных и агрессивных (рис. 5). Процент выборов во время латеральных предпочтений (предпочтений пустой миски, предъявляемой на одной стороне) в этом тесте оказался также достоверно ниже у неселекционируемых лисиц, чем у ручных и агрессивных. В то время как между выборками из контрастных по поведению популяций не было показано достоверных различий по этому показателю (рис. 5).
Рис. 5.
Процент выборов лисицами пустой миски (“0”) и процент выборов во время латеральных предпочтений (“Л”); тест 4:0, описание в тексте. ** – p < 0.01 по сравнению с ручными и агрессивными лисицами.
Fig. 5. The percent of choices of empty container (“0”) and percent of choices during uni-lateral choices (“Л”); test 4:0, description in the text. ** – p < 0.01 in comparison with tame and aggressive foxes.
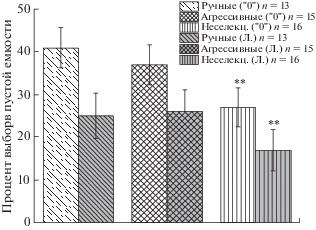
В эксперименте с равным количеством кусочков печени (4:4) мы оценивали процент выборов во время латеральных предпочтений. Если лисица два или более раз подряд выбирала емкость с кормом на одной стороне (независимо – справа или слева), эти выборы мы относили к латеральным предпочтениям. Данный показатель в этом тесте во всех трех исследованных группах оказался достоверно выше случайного уровня (50%). Он составил для ручных лисиц 84,9% выборов (n = 14; p ≤ ≤ 0.01 по сравнению со случайным уровнем 50:50, F-критерий), для агрессивных лисиц 86% выборов (n = 15; p ≤ 0.01 по сравнению со случайным уровнем 50:50, F-критерий), для неселекционируемых по поведению лисиц 86,8% выборов (n = 16; p ≤ 0.01 по сравнению со случайным уровнем 50:50, F-критерий). Между представителями из популяций по данному показателю не было показано достоверных различий.
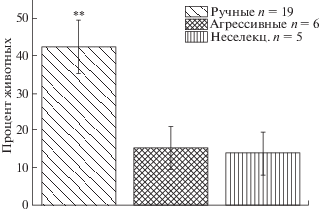
Также мы считаем интересными данные о доле лисиц (рис. 6), которых пришлось исключить из эксперимента во время первого дня тестирования (когда предъявлялось соотношение 10 и 1 кусочков). Эти животные сначала проявляли безудержные попытки утащить одну из емкостей с кормом в глубь клетки без стремления съесть корм, затем после естественного угасания этой реакции сильного возбуждения они уходили к дальней стенке клетки и переставали подходить к мискам с кусочками корма. Если животное не подходило к емкостям с кормом в течение пяти минут, экспериментатор принимал решение исключить его из эксперимента. Такое поведение наблюдалось либо на самом последнем этапе адаптации к процедуре тестирования (видеозапись доступна на сайте: http://www.bionet.nsc.ru/razrabotki/prikladnyie-razrabotki/zhivotnovodstvo/lisiczyi/ruchnaya-lisicza-vozbuzhdenie-i-posleduyushhij-otkaz-ot-korma.html), когда экспериментатор предъявлял лисице одну из емкостей с кусочком пищи, дожидаясь, пока она не начинала съедать корм сразу при помещении емкости в прорезь клетки. Либо лисица могла демонстрировать такое поведение в любой момент в течение первого теста (10:1), успешно пройдя все предыдущие этапы адаптации к нему. На рисунке 6 видно, что доля животных, которых пришлось исключить, оказалась достоверно выше у ручных лисиц, чем у агрессивных и неселекционируемых.
Рис. 6.
Процент лисиц, которые были исключены из эксперимента во время первого теста (10:1). ** p ≤ 0.01 по сравнению с агрессивными и неселекционируемыми лисицами.
Fig. 6. Percent of foxes, who was excluded from experiment in test 10:1. ** p ≤ 0.01 in comparison with aggressive and unselected foxes.
ОБСУЖДЕНИЕ РЕЗУЛЬТАТОВ
На основе полученных результатов, несмотря на большие индивидуальные различия между лисицами в решении тестов, мы попытались выделить для каждой из исследованных групп некоторые общие характеристики поведения.
Доместицируемые лисицы чаще меняют сторону выбора, не проявляя латеральных предпочтений какой-либо одной стороны (рис. 1), что, по нашему мнению, характеризует большую пластичность их поведения по сравнению с агрессивными и неселекционируемыми, которая была также показана ранее в тесте на переделку инструментального навыка [Мухамедшина и др., 2019]. Именно предъявление большего количества на одной и той же стороне при такой стратегии поведения делает возможным более быстрое обучение методом проб и ошибок (видеозапись доступна на сайте: http://www.bionet.nsc.ru/razrabotki/prikladnyie-razrabotki/zhivotnovodstvo/lisiczyi/ruchnaya-lisicza-obuchaetsya-vyibirat-bolshee-kolichestvo-kusochkov-metodom-prob-i-oshibok.html). Все лисицы, успешно выбиравшие большее количество, демонстрировали низкий уровень односторонних выборов (рис. 3). Доля односторонних выборов у ручных ниже, чем у агрессивных и неселекционируемых лисиц, что, по нашему мнению, влияет на их успешность в выборе большего количества в задаче 5:1 (рис. 2). Тем не менее с возрастанием сложности задачи ручные также начинают чаще выбирать емкость с кормом на одной и той же стороне (рис. 1).
В экспериментах по оценке способности к экстраполяции направления движения стимула Л.В. Крушинский также наблюдал аналогичную форму поведения у разных видов животных – односторонние обходы ширмы, не зависящие от направления движения стимула. Одной из причин односторонних обходов он предполагал перенапряжение нервной системы после неоднократного предъявления сложной задачи как своеобразную форму защиты ЦНС от перенапряжения [Крушинский, 2009]. Он наблюдал подобные ухудшение решения задач и невротические состояния только у успешно решающих особей. Гипотеза о том, что одной из причин, по которой животные могут предпочтительно выбирать емкость с одной и той же стороны, является нервная перегрузка, подтверждается и результатами дополнительного теста, в котором одна из емкостей, вдвигаемых экспериментатором в домашнюю клетку животного, оставалась пустой (рис. 4).
Процент выборов пустой миски у ручных лисиц не отличался от такового у агрессивных, но был достоверно выше, чем у неселекционируемых лисиц (при этом среди всех протестированных лисиц мы не обнаружили ни одной, которая выбирала бы пустую миску на протяжении всего теста). Хронологически этот тест проводился нами в последнюю очередь. Такое на первый взгляд ненормальное поведение ручных лисиц можно объяснить, по-видимому, их неспособностью затормозить поведение, которое ранее приводило к получению пищи, поскольку во всех предыдущих тестах пища находилась в обеих мисках. Это подтверждается результатами теста на способность к угашению, которая также оказалась ниже у ручных [Мухамедшина и др., 2014].
Что касается агрессивных лисиц, некоторые из них во время теста 4:0, при выборе пустой емкости в состоянии возбуждения начинали демонстрировать агрессивное поведение по отношению к вдвигаемой емкости, но при следующем предъявлении они снова выбирали пустую емкость (видеозапись доступна на сайте: http://www.bionet.nsc.ru/razrabotki/prikladnyie-razrabotki/zhivotnovodstvo/lisiczyi/agressivnaya-lisicza-demonstriruet-predpochtenie-pustoj-emkosti-v-dopolnitelnom-eksperimente.html). Такое поведение является ярким примером того, что в случаях выборов пустой миски из-за высокого напряжения нервной системы агрессивные лисицы так же, как и ручные, не могут затормозить эту форму поведения, но, с нашей точки зрения, по другой причине. В тесте 4:0 перегрузка у агрессивных лисиц, по-видимому, вызывается необходимостью делать выбор, что может являться подтверждением снижения у них подвижности нервных процессов. О снижении подвижности нервных процессов у агрессивных лисиц свидетельствует также и более низкая скорость переделки инструментального навыка [Мухамедшина и др., 2019].
В тесте 4:0 неселекционируемые лисицы демонстрируют достоверно меньший процент латеральных предпочтений и выборов пустой миски, чем ручные и агрессивные лисицы. По-видимому, они в целом менее эмоциональны и меньше подвержены фрустрации, чем ручные и агрессивные, поэтому способны затормозить выбор пустой миски, который не является адаптивным. Меньшая эмоциональность неселекционируемых лисиц подтверждается и данными по их вокальной активности – частота следования звуков (которая является косвенным показателем повышенной эмоциональности), у них ниже по сравнению с ручными и агрессивными [Гоголева и др., 2017], а также нашими исследованиями [Мухамедшина и др., 2019].
Неселекционируемые по поведению лисицы явились исходной популяцией, с которой начинался процесс отбора как на ручное, так и на агрессивное поведение. Однако фактически неселекционируемые лисицы также в некоторой степени претерпевают изменения в сторону повышения толерантности по отношению к человеку. Но в целом по многим показателям эта популяция оказывается ближе к агрессивным лисицам. Так, было показано, что неселекционируемые и агрессивные лисицы используют одинаковый набор звуков при контактах с человеком [Гоголева и др, 2017]. По содержанию АКТГ и кортизола в плазме крови не было различий между агрессивными и неселекционируемыми по поведению лисицами ни в покое, ни на фоне эмоционального стресса [Оськина и др., 2008]. По проценту выборов большего количества кусочков и проценту выборов во время латеральных предпочтений неселекционируемые лисицы также оказались ближе к агрессивным лисицам, чем к ручным. Врожденная осторожность лисиц из неселекционируемой популяции ограничивает их исследовательское поведение. Поэтому нам представляется естественным, что неселекционируемые лисицы не стремятся к активному поиску вариантов при решении когнитивной задачи. Вероятно, это может являться одной из причин, по которой неселекционируемые лисицы оказались склонными к односторонним выборам.
Из представленных в нашей работе данных следует, что лисица (Vulpes vulpes) демонстрирует достаточно большую вариабельность реакций в предъявленных тестах, которая оказалась во многом связанной с характером поведения по отношению к человеку.
ВЫВОДЫ
1. Основное различие в стратегии поведения лисиц в тестах с предъявлением разного количества кусочков корма по заданной схеме сводится к большей выраженности у агрессивных и неселекционируемых такой формы поведения, как выбор емкости с какой-либо одной стороны (либо только правой, либо левой), независимо от количества кусочков пищи в ней.
2. При решении наиболее простой задачи (различение 10 и 1 кусочков корма) одинаковый процент лисиц из ручной, агрессивной и неселекционированной по поведению популяций достигли критерия неслучайности выборов большего количества кусочков пищи.
3. Ручные лисицы продемонстрировали большую успешность в решении задачи 5 и 1 по сравнению с агрессивными и неселекционируемыми, благодаря большей вариативности их поведения (более частая смена сторон выбора).
Выражаем благодарность сотрудникам ЦКП “Генофонды пушных и с/х животных” ИЦиГ СО РАН за помощь в работе с животными, ЦКП “Генофонды пушных и с/х животных” ИЦиГ СО РАН за предоставленных животных, а также В.М. Ефимову за консультирование по вопросам статистического анализа результатов. Работа по тестированию животных выполнена при поддержке гранта РНФ № 16-14-10216. Финансирование инфраструктуры для исследований выполнено за счет бюджетного проекта № 0324-2018-0016.
Список литературы
Беляев Д.К. Дестабилизирующий отбор. Развитие эволюционной теории в СССР (1917–1970-е годы). Отв. ред. Микулинский С.Р., Полянский Ю.И. Л.: Наука, 1983. 266–277.
Васильева Л.Л. Феногенетический анализ поведения серебристо-черных лисиц (Vulpes vulpes) при ослаблении эффективности отбора на доместикацию. Дисс. … к.б.н. Новосибирск. ИЦиГ СО РАН. 1991а. 184 с.
Васильева Л.Л. Анализ эффекта доместикации в изменении способности серебристо-черных лисиц (Vulpes vulpes) к обучению. В Сб.: Эволюционно-генетические и генетико-физиологические аспекты доместикации пушных зверей. Новосибирск. 1991б. с. 57–69.
Гоголева С.С., Володин И.А., Володина Е.В., Харламова А.В., Трут Л.Н. Влияние экспериментальной доместикации серебристо-черных лисиц (Vulpes vulpes) на вокальное поведение. Вавиловский журнал генетики и селекции. 2017. 21 (4): 402–413.
Зорина З.А., Смирнова А.А. Относительные количественные оценки у голубей и ворон: спонтанный выбор большего пищевого множества Журн. высш. нерв. деят. 1994. 44 (3): 618–621.
Зорина З.А., Смирнова А.А. Количественная оценка серой вороной множеств, состоящих из 15-25 элементов. Журн. высш. нерв. деят. 1996. 46 (2): 298–301.
Крушинский Л.В. Биологические основы рассудочной деятельности: Эволюционный и физиолого-генетический аспекты поведения. М.: Книжный дом “Либроком”, 2009. 272 с.
Лоренц К. Оборотная сторона зеркала. М.: Республика, 1998. 393 с.
Мухамедшина И.А., Харламова А.В., Трут Л.Н. Изменяет ли отбор лисиц на доместикацию и агрессивность их способность концентрировать внимание и формировать двигательный навык? Журн. высш. нерв. деят. 2014. 64 (5): 521–530.
Мухамедшина И.А., Харламова А.В., Трут Л.Н. Некоторые особенности высшей нервной деятельности лисиц и влияние на них отбора по социальным реакциям на человека. Журн. высш. нерв. деят. 2019. 69 (1):88–97.
Оськина И.Н., Гербек Ю.Э., Шихевич С.Г. Плюснина И.З., Гулевич Р.Г. Изменения гипоталамо-гипофизарно-надпочечниковой системы при отборе животных на доместикационное поведение. Вестн. ВОГиС. 2008. 2 (1/2): 39–49.
Плескачева М.Г. Поведение и пространственное обучение птиц в радиальном лабиринте. Журн. высш. нерв. деят. 2008. 58 (4): 389–407.
Резникова Ж.И. Экспериментальные исследования способностей животных к количественным оценкам предметного мира. Модернизм в психологии. Сб. мат-лов Всеросс. конф. Новосибирск: Изд-во НГУ, 2004. 113–136.
Трут Л.Н. Роль поведения в доместикационных преобразованиях животных (на примере серебристо-черных лисиц). Дисс. … д.б.н. Новосибирск, 1980. 410 с.
Banerjee1 K., Chabris C., Johnson V., Lee1 J., Tsao F., Hauser M. General Intelligence in Another Primate: Individual Differences across Cognitive Task Performance in a New World Monkey (Saguinus oedipus). PLoS ONE. 2009. 4 (6). e5883.
Banszegi O., Urrutia A., Szenczi P., Hudson R. More or less: spontaneous quantity discrimination in the domestic cat. Anim. Cogn. 2016. https://doi.org/10.1007/s10071-016-0985-2
Benjamini Y., Hochberg Y. Controlling the false discovery rate: A practical and powerful approach to multiple testing. Journal of the Royal Statistical Society, Series B. 1995. 57: 289–300.
Broadway M., Samuelson M., Christopher J., Jett S., Lyn H. Does size really matter? Investigating cognitive differences in spatial memory ability based on size in domestic dogs. Behav Processes. 2017. https://doi.org/10.1016/j.beproc.2017.01.012
Brucks D., Marshall-Pescini S., Wallis L., Huber L., Range F. Measures of Dogs’ Inhibitory Control Abilities Do Not Correlate across Tasks. Front. Psychol. 2017. 8:849. https://doi.org/10.3389/fpsyg.2017.00849
Gallistel C., Gelman R. Non verbal numerical cognition: from reals to integers. Trends in Cognitive science. 2000. 4 (2): 59–65.
Hanus D., Call J. Discret equantity judgments in the great apes (Pan paniscus, Pan troglodytes, Gorilla gorilla, Pongo pygmaeus): the effect of presenting whole sets versus item-by-item. J. Comp. Psychol. 2007. 121: 241–249.
Hunt S., Low J., Burns K. Adaptive numerical competency in a food-hoarding songbird. Proc Biol Sci. 2008. 275: 2373–2379.
Jo Y., Lee I. Perirhinal cortex is necessary for acquiring, but not for retrieving object-place paired association. Learn. Mem. 2010. 17: 97–103.
Lee I., Kim J. The shift from a response strategy to object-in-place strategy during learning is accompanied by a matching shift in neural firing correlates in the hippocampus. Learn. Mem. 2010. 17: 381–393.
Lee I., Solivan F. The roles of the medial prefrontal cortex and hippocampus in a spatial paired-association task. Learn. Mem. 2008. 15: 357–367.
Lee I., Solivan F. Dentate gyrus is necessary for disambiguating similar object-place representations. Learn. Mem. 2010. 17: 252–258.
Lewis K., Jaffe S., Brannon E. Analog number representations in mongoose lemurs (Eulemur mongoz): evidence from a search task. Anim. Cogn. 2005. 8 (4): 247–252.
Lipp H.P., Pleskacheva M.G., Gossweiler H. et al. A large outdoor radial maze for comparative study in birds and mammals. Neurosci. Biobehav. Rev. 2001. 25 (1): 83–99.
MacLean E., Hare B., Nunn C., Addessi E., Amici F., Anderson R., Aureli F., Baker J., Bania A., Barnard A., Boogert N., Brannon E., Bray E., Bray J., Brent L., Burkart J., Call J., Cantlon J., Cheke L., Clayton N., Delgado M., DiVincenti L., Fujita K., Herrmann E., Hiramatsu C., Jacobs L., Jordan K., Laude J., Leimgruber K., Messer E., Moura A., Ostojić L., Picard A., Platt M., Plotnik J., Range F., Reader S., Reddy R., Sandel A., Santos L., Schumann K., Seed A., Sewall K., Shaw R., Slocombe K., Su Y., Takimoto A., Tan J., Tao R., van Schaik C., Virányi Z., Visalberghi E., Wade J., Watanabe A., Widness J., Young J., Zentall T., Zhao Y. The evolution of self-control. Proc Natl Acad Sci USA. 2014. 111 (20). https://doi.org/10.1073/pnas.1323533111
Macpherson K., Roberts W. Can dogs count? Learning and Motivation. 2013. https://doi.org/10.1016/j.lmot.2013.04.002
Osthaus B., Marlow D., Ducat P. Minding the gap: spatial perseveration error in dogs. Anim. Cogn. 2010. 13: 881–885.
Osthaus B., Proops L., Hocking I., Burden F. Spatial cognition and perseveration by horses, donkeys and mules in a simple A-not-B detour task. Anim. Cogn. 2013. 16: 301–305.
Petrazzini M., Wynne C. What counts for dogs (Canis lupus familiaris) in a quantity discrimination task? Behav. Proc. 2016. 122. 90–97.
Petrazzini M., Wynne C. Quantity discrimination in canids: dogs (Canis familiaris) and wolves (Canis lupus) compared. Behav. Proc. 2017. https://doi.org/10.1016/j.beproc.2017.09.003
Range F., Jenikejew J., Schröder I., Virányi Z. Difference in quantity discrimination in dogs and wolves. Froint. Psychol. 2014. 18.
Reznikova Zh., Ryabko B. Numerical competence in animals, with an insight from ants. Behaviour 2011. 148: 405–434.
Topál J., Gergely G., Erdohegyi A., Csibra G., Miklósi A. Differential sensitivity to human communication in dogs, wolves, and human infants. Science. 2009. 325: 1269–1272.
Trut L.N. Early canid domestication: farm-fox experiment. Amer. Sci. 1999. 87: 160–169.
Trut L., Oskina I., Kharlamova A. Animal evolution during domestication: the domesticated fox as a model. BioEssays. 2009. 31: 349–360.
Tryon R.C. Studies in individual differences in maze ability. VII. The specific components of maze ability and general theory of psychological components. – J. Comp. Physiol. Psycol. 1940. 30 (2): 283–335.
Ward C., Smuts B.B. Quantity-based judgments in the domestic dog (Canis lupus familiaris). Anim. Cogn. 2007. 10 (1): 71–80.
Дополнительные материалы отсутствуют.
Инструменты
Журнал высшей нервной деятельности им. И.П. Павлова