

Журнал высшей нервной деятельности им. И.П. Павлова, 2019, T. 69, № 5, стр. 549-560
ПОКАЗАТЕЛИ ДИФФУЗИИ ТРАКТОВ ГОЛОВНОГО МОЗГА, ФУНКЦИОНАЛЬНАЯ СВЯЗАННОСТЬ ВО ФРОНТО-ПАРИЕТАЛЬНОЙ СЕТИ И ПАРАМЕТРЫ УПРАВЛЯЮЩИХ ФУНКЦИЙ У МОЛОДЫХ ЗДОРОВЫХ ИСПЫТУЕМЫХ
И. В. Клочкова 1, *, Я. Р. Паникратова 1, М. В. Алфимова 1, А. С. Томышев 1, Д. А. Куприянов 2, И. С. Лебедева 1
1 Лаборатория нейровизуализации и мультимодального анализа, ФГБНУ НЦПЗ
Москва, Россия
2 ООО “Филипс”
Москва, Россия
* E-mail: klochkova1804@list.ru
Поступила в редакцию 28.12.2017
После доработки 21.12.2018
Принята к публикации 11.02.2019
Аннотация
Целью настоящего исследования был поиск взаимосвязей между параметрами управляющих функций и характеристиками их структурного (особенности трактов головного мозга) и функционального (фронто-париетальная нейрональная сеть) обеспечения у молодых здоровых испытуемых с использованием мультимодального подхода, сочетающего диффузионно-взвешенную МРТ, функциональную МРТ покоя и нейропсихологические пробы. В исследовании приняли участие 16 праворуких испытуемых, 8 мужчин и 8 женщин (19–32 года). Нейропсихологическое обследование проводили с помощью тестов батареи Delis–Kaplan Executive Function System; структурная, диффузионно-взвешенная и функциональная МРТ покоя проводились на томографе Philips 3T (Голландия). Сопоставление данных фМРТ покоя и нейропсихологических данных показало, что слабость переключения ассоциировалась с меньшей связанностью между височным и поясным компонентами фронто-париетальной сети в правом полушарии. Корреляции между трактографическими и нейропсихологическими характеристиками не прошли коррекцию на множественность сравнений: между радиальной диффузией передней таламической лучистости и относительно простой когнитивной характеристикой – скоростью называния цветов в тесте Струпа. Полученные результаты расширяют представления об ассоциации структурной и функциональной связанности головного мозга с конкретными компонентами управляющих функций у молодых здоровых испытуемых.
ВВЕДЕНИЕ
Одним из ключевых элементов обеспечения психической жизни человека являются т.н. управляющие функции (УФ, executive functions), объединяющие процессы избирательной регуляции, программирования, планирования и контроля целенаправленного поведения [Мачинская, 2015].
Проблема нейрофизиологической организации и нейробиологического базиса когнитивных функций, в том числе и управляющих, как наиболее сложно организованных и уязвимых, является одной из наиболее исследуемых и тем не менее на данный момент не решенных проблем. Специфика исследуемой области заключается в сложной иерархии структурно-функционального обеспечения всех когнитивных функций как высших психических функций [Лурия, 1973], что не позволяет дать однозначный ответ об их мозговой организации. Основываясь на исследованиях локальных поражений мозга, УФ традиционно связывали с функционированием лобных долей головного мозга (термины “УФ” и “лобные функции” даже использовались как синонимы) [Лурия, 1962; Stuss, 2011].
С возникновением и развитием современных методов нейровизуализации открылся широкий круг возможностей исследования когнитивных функций, появились новые данные относительно их организации.
Исследования с применением функциональной МРТ (фМРТ), связанной с выполнением задачи, продемонстрировали, что ключевой функциональной сетью, участвующей в обеспечении УФ, является фронто-париетальная нейрональная сеть [метаанализы Niendam et al., 2012; Collette et al., 2005; Sylvester et al., 2003; обзоры Zanto, Gazzaley, 2013; Ptak, 2011; Friedman, Miyake, 2017]. Ее основные компоненты располагаются в дорсолатеральной префронтальной коре, медиальной префронтальной коре (в частности, в передней поясной извилине) и теменных отделах [Niendam et al., 2012; Collette et al., 2005; Sylvester et al., 2003], однако выделяют также компоненты в нижних височных отделах [Yeo et al., 2011]. Метод фМРТ покоя, позволяющий исследовать функциональную организацию мозга при отсутствии внешней задачи, также позволяет выделить эту сеть [Mazoyer et al., 2001; Yeo et al., 2011].
Обращает на себя внимание немногочисленность исследований на здоровой выборке молодого возраста, которые бы совмещали нейропсихологические методы с фМРТ покоя для исследования связей УФ с функционированием фронто-париетальной сети. Однако данное направление исследований является важным как для изучения мозговой организации УФ в норме, так и для дальнейшего анализа патологий в функционировании данной сети при психиатрических и неврологических заболеваниях. В исследовании Seeley и коллег [2007] усиление функциональной связанности (ФС) между двумя компонентами фронто-париетальной сети (во внутритеменной борозде/верхней теменной дольке билатерально) коррелировало с высокими показателями такого компонента УФ, как переключение (измерялось как разница между временем выполнения субтестов А и B теста “Trail Making Test”). Reineberg и коллеги [2016] изучали связь индивидуальных различий в 3 компонентах УФ (Общая УФ, переключение и рабочая память) с внутренней и внешней функциональной связанностью (ФС) фронто-париетальной сети. Более высокие показатели по общей УФ оказались связаны с повышением ФС клина и дополнительной моторной коры и со снижением ФС в латеральных лобных отделах, а также левой височной зоне; более высокие показатели рабочей памяти – со снижением ФС латеральных и медиальных компонентов фронто-париетальной сети. Коррелятов для переключения в рамках фронто-париетальной сети этими авторами выявлено не было. Chen и коллеги [2018] провели метаанализ, позволивший выделить общие зоны активации как для эмоционального, так и для классического теста Струпа (направленного на оценку оттормаживания): передняя островковая кора и дорсальная передняя поясная кора/дополнительная моторная кора билатерально, нижняя лобная извилина и верхняя теменная долька в левом полушарии. В состоянии покоя эти области демонстрируют ФС с одними и теми же зонами мозга, образуя сеть, включающую дополнительно области в средней и медиальной лобных извилинах, прецентральной извилине, средней височной извилине, таламусе, мозжечке и базальных ганглиях (эти области относились преимущественно к фронто-париетальной сети). В исследовании Takeuchi и коллег [2015] было показано, что более успешное выполнение теста Струпа ассоциировалось с большим индексом центральности (degree centrality; показатель ФС между зоной интереса и остальными зонами мозга) в левой латеральной префронтальной коре, нижней лобной извилине и нижней теменной дольке левого полушария. Учитывая небольшое число работ, посвященных ассоциации показателей УФ и ФС внутри фронто-париетальной сети в состоянии покоя, а также разнородность результатов, мы исследовали корреляции между нейропсихологическими показателями УФ и ФС внутри фронто-париетальной сети.
Относительно связи управляющих функций со структурными характеристиками головного мозга полученные к настоящему моменту данные указывают, что ключевой локус “мозгового обеспечения” УФ находится в лобных долях, в первую очередь, дорсолатеральной префронтальной коре (ДЛПК) [Мачинская, 2015; Alvarez, Emory, 2006; Nowrangi et al., 2014; Gilbert, Burgess, 2008], хотя в эти процессы “включены” и многие другие области головного мозга: теменные отделы [Garavan et al., 2002; Szameitat et al., 2002; Petersen, Posner, 2012; Chung et al., 2014], базальные ганглии [Monchi et al., 2006], таламус [Chung et al., 2014; Мачинская, 2001], мозжечок [Bellebaum, Daum, 2007]. Данные факты отвечают представлениям о распределенности анатомо-морфологического обеспечения когнитивных функций [Mesulam, 1990; Zeki, Shipp, 1988], их иерархии как интеграционных образований локальных специализированных процессов обработки информации [Park and Friston, 2013].
Как следствие, одним из направлений исследований стал анализ так называемой структурной связанности ДЛПК и других отделов головного мозга, а объективным, информативным и неинвазивным методом ее изучения – диффузионно-взвешенная томография (ДВ-МРТ).
К настоящему времени получен достаточно большой массив данных, характеризующихся, однако, определенной гетерогенностью. Так, например, было выявлено, что большая структурированность белого вещества в областях, подлежащих префронтальной коре, связана с более высокими показателями УФ [Seghete et al., 2013], а более высокая фракционная анизотропия (лучшая структрированность) трактов, соединяющих ветромедиальные и вентролатеральные области с каудальными областями головного мозга, ассоциировалась с лучшим выполнением тестов на УФ [Smolker et al., 2015].
В одном из исследований авторы указывали на существование обширных областей (от префронтальной к теменной коре), для которых наблюдалась максимальная корреляция между характеристиками УФ и микроструктурными характеристиками белого вещества [Grieve et al., 2007]. В других исследованиях определяющей оказалась вариабельность структуры крючковидного и верхнего продольного пучков [Sasson et al., 2013], правого крючковидного и правого поясного пучков [Lebel et al., 2013] или правого предклинья [Takeuchi et al., 2011].
С другой стороны, Vollmer et al. [2017] обнаружили прямые корреляции между характеристиками рабочей памяти и фракционной анизотропией передней таламической лучистости, крючковидного пучка, нижнего продольного пучка и нижнего лобно-затылочного пучка, но только у подростков, родившихся недоношенными.
Целью настоящего исследования стало выявление взаимосвязей между параметрами структурной и функциональной связанности зон головного мозга и характеристиками управляющих функций. Соответственно цели была поставлена задача данного фрагмента исследования – определение корреляций между параметрами структурной связанности (определяемой по ДВ-МРТ), показателями функциональной связанности фронто-париетальной сети (определяемой по фМРТ покоя) и характеристиками управляющих функций (определяемыми по нейропсихологическим тестам батареи Delis–Kaplan Executive Function System). Исходя из данных литературы, было выдвинуто предположение о том, что вариабельность в структурно-функциональных показателях головного мозга будет коррелировать с тестируемыми характеристиками управляющих функций. Исследование проводилось на испытуемых молодого возраста, что крайне важно для развития представлений о связях структура-функция в целом, а также создает фундамент для изучения изменения этих связей при психических заболеваниях, начинающихся в молодом и среднем возрасте, таких, например, как шизофрения или биполярное расстройство.
Особенностью настоящей работы стало применение метода вероятностной автоматической трактографии TRACULA (TRActs Constrained by UnderLying Anatomy) [Yendiki et al., 2011], что и определило новизну и актуальность работы. Данный алгоритм использует не только диффузионно-взвешенные изображения, но и анатомические реконструкции головного мозга каждого испытуемого (полученные по данным структурной МРТ), а также информацию о вероятностях прохождения трактов через (или около) различные анатомические структуры, тем самым позволяя автоматически реконструировать проводящие пути в индивидуальном пространстве головного мозга каждого испытуемого. Помимо этого, TRACULA использует модель диффузии “ball-and-stick”, обладающую большей точностью в вокселях, включающих пересекающиеся проводящие пути, по сравнению с традиционной тензорной моделью [Behrens et al., 2007].
Дополнительно в данной работе был проведен анализ с использованием нейровизуализационного показателя функциональной связанности – с помощью метода фМРТ покоя была выделена так называемая фронто-париетальная нейрональная сеть, описываемая как ключевая при обеспечении УФ [обзоры Zanto, Gazzaley, 2013; Ptak, 2011; Friedman, Miyake, 2017]. В связи с ограничениями метода фМРТ, связанными с выполнением задачи (высокая зависимость результатов от конкретной выбранной задачи и особенностей предъявления стимульного материала), в последнее время наиболее широко применяется фМРТ покоя, при которой регистрация данных проводится в отсутствие “внешней” задачи. Применение фМРТ покоя позволяет увидеть общие особенности функционирования мозга в норме и патологии (функциональные профили), в отличие от частной ситуации выявления активации определенных областей мозга при выполнении конкретной задачи.
МЕТОДИКА
Объект исследования
Исследование было проведено в ФГБНУ НЦПЗ. В исследовании приняли участие 16 испытуемых, 8 мужчин и 8 женщин, средний возраст по выборке составил 23.68 ± 4.03 г. (19–32 года). Критериями включения были: праворукость, отсутствие в анамнезе психических или тяжелых неврологических заболеваний, зависимостей, отсутствие нарушений слуха, отсутствие психически больных родственников первой и второй степени родства.
Испытуемые имели следующие уровни образования: неоконченное высшее образование (студенты 1–4 курса вузов) – 8 человек, высшее образование – 7 человек, более одного высшего образования – 1 человек. Все испытуемые на момент обследования были соматически здоровы, не имели противопоказаний к прохождению всех этапов исследования.
Исследование проводилось в соответствии с этическими принципами проведения медицинских исследований с участием людей в качестве субъектов (Хельсинкская декларация Всемирной медицинской ассоциации). Все испытуемые подписывали добровольное информированное согласие на участие в обследовании.
Нейропсихологическое обследование
Обследование проводилось с использованием следующих тестов батареи Delis–Kaplan Executive Function System [Delis, Kaplan, Kramer, 2004]: модификация теста Струпа (Color-Word Interference task), тест Визуальной беглости (Design fluency test), тест Вербальной беглости (Verbal fluency test). По тесту Струпа оценивали показатели времени выполнения серии 1 (называние цветов), серии 2 (чтение слов), серии 3 (называние цвета чернил, которым были напечатаны названия цветов, в конфликтных условиях – при несовпадении значения слова и цвета чернил), и серии 4 (переключение с называния цвета на чтение слова в конфликтных условиях), а также разность времени серий 3 и 1 (эффект Струпа, или оттормаживание) и серий 4 и 3 (эффект переключения, или когнитивная гибкость); по тесту визуальной беглости – количество правильных ответов по сериям 1 (создание фигур путем соединения черных точек), 2 (из белых точек, игнорируя черные), 3 (чередуя белые и черные точки); по тесту вербальной беглости – количество правильных ответов по сериям 1 (фонетическая вербальная беглость), 2 (семантическая вербальная беглость), 3 (семантическая вербальная беглость с переключением). Каждый испытуемый выполнял все серии указанных тестов, результаты фиксировались в протоколе ответов.
Исследование проводили в тот же день, что и МРТ, максимально приближенно по времени к томографическому обследованию.
МРТ-исследование
МРТ-исследование (в рамках тестирования стандартных программ) проводилось на МР томографе 3Т Philips (Голландия). На первом этапе были получены анатомические T1-изображения (170 срезов, размер воксела 1 × 1 × 1 мм, 3DTFE (TR/TE/FA = = 8 мс/3.5 мс/8°)). Функциональные T2*-изображения фМРТ покоя были получены с помощью последовательности EPI (TR/TE/FA = = 3 с/35 мс/90°), 35 срезов, матрица 128 × 128 вокселов, размер воксела 1.8 × 1.8 × 4 мм. Во время фМРТ-сканирования испытуемые лежали неподвижно с открытыми глазами, фиксируя взгляд на белом кресте в центре серого поля монитора в течение 5 мин. Испытуемым была дана инструкция избегать любой систематической мыслительной деятельности.
Диффузионно-взвешенные изображения были получены во время того же обследования с использованием последовательности спин-эхо EPI (TR/TE/FA = 4075 мс/87 мс/90°; поле обзора = 240 мм). Диффузионные градиенты были приложены в 32 неколлинеарных направлениях с фактором диффузии b = = 800 с/мм2 и b = 100 с/мм2 (во время анализа второй b-фактор был исключен), и в одном направлении с фактором диффузии b = 0.
Обработка данных – диффузионно-взвешенная томография
Реконструкция коры и сегментация подкоркового вещества по данным структурной МРТ проводилась с использованием пакета FreeSurfer 5.3.0 (MGH/MIT/Harvard Medical School, Athinoula A. Martinos Center for Biomedical Imaging). Далее, используя анатомические реконструкции, полученные во FreeSurfer 5.3.0, и модель диффузии, построенную с помощью инструмента BEDPOSTX (часть пакета FSL 5.0.7, Oxford Centre for Functional Magnetic Resonance Imaging of the Brain Software Library), при помощи алгоритмов TRACULA (часть FreeSurfer 5.3.0) была проведена автоматическая (машинная) реконструкция восемнадцати трактов белого вещества для каждого испытуемого (рис. 1).
Рис. 1.
Реконструкция 18 трактов одного из испытуемых. Тракты изображены в виде изоповерхностей на фоне T1-взвешенного изображения этого же пациента. Аксиальная и саггитальная проекции.
Fig. 1. Reconstruction of 18 tracts of one of the subjects. The tracts are depicted as isosurfaces on the background of a T1-weighted image of the same patient. Axial and sagittal projection.
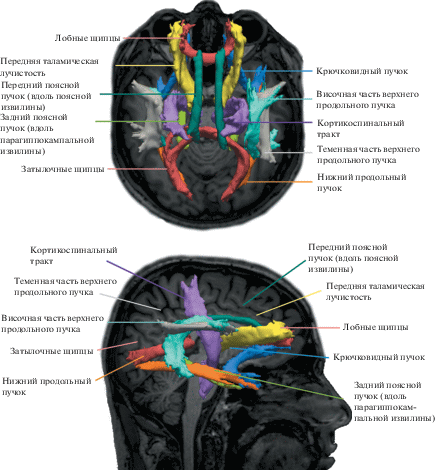
Для анализа были использованы средневзвешенные по всему тракту показатели фракционной анизотропии (FA), аксиальной (AD) и радиальной диффузии (RD).
Обработка данных – фМРТ покоя
Предобработку изображений проводили с использованием пакета SPM12 (http://www.fil.ion.ucl.ac.uk/spm/software/spm12/). Изображения ориентировали параллельно плоскости, проходящей через переднюю и заднюю комиссуры (AC/PC). Производились коррекция метрических искажений, возникающих вследствие неоднородности магнитного поля, и его взаимодействия с движением головы испытуемого (realign & unwarp), корректировка смещения во времени измерений в рамках одного объема головного мозга, совмещение функциональных изображений с анатомическими, выделение в анатомических изображениях серого, белого вещества и спинномозговой жидкости, приведение всех изображений к координатам пространства MNI (при этом размер воксела анатомических изображений остался прежним, размер воксела функциональных изображений был изменен на 3 × 3 × 3 мм), пространственное сглаживание функциональных изображений (8 мм).
Дальнейшая обработка данных фМРТ покоя проводилась с помощью CONN-fMRI toolbox 17.b [Whitfield-Gabrieli, Nieto-Castanon, 2012]. Коррекция артефактов движения включала снижение веса в анализе измерений, во время которых регистрировалось движение головы испытуемых (“ART-based identification of outlier scans for scrubbing” с пороговыми значениями 97 перцентилей сохранных изображений в нормативной выборке). Кроме того, на этапе шумоподавления выделялись вокселы, флуктуации BOLD-сигнала в которых коррелировали с колебаниями сигнала в белом веществе или спинномозговой жидкости (по индивидуальным картам). Производился учет индивидуальных параметров движения как ковариат первого уровня. Применялся частотный фильтр (0.008 < f < 0.09 Гц).
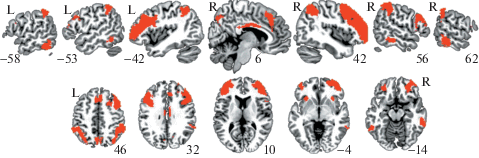
Данные фМРТ покоя обрабатывались методом анализа функциональных связей между зонами интереса (regions of interest; “ROI-to-ROI analysis”) в состоянии покоя испытуемого. Связи оценивались с помощью коэффициентов корреляции с трансформацией Фишера между каждой парой зон. Зонами интереса выступили компоненты фронто-париетальной сети (всего 16 компонентов по обоим полушариям; таблица 1), выделенные методом анализа функциональных связей в состоянии покоя на 1000 испытуемых [Yeo et al., 2011, см. рис. 2, подробное описание см., например, https://surfer.nmr.mgh.harvard.edu/fswiki/CorticalParcellation_Yeo2011)].
Таблица 1.
Анатомическое расположение компонентов фронто-париетальной сети атласа Yeo et al. [2011] Table 1. Anatomical localization of the fronto-parietal network components of the atlas by Yeo et al. [2011]
| Левое полушарие | Правое полушарие | ||
|---|---|---|---|
| 1 | Теменные отделы | 10 | Теменные отделы |
| 2 | Височные отделы | 11 | Височные отделы |
| 3 | Дорсальная префронтальная кора | ||
| 4 | Латеральная префронтальная кора | 12 | Латеральная префронтальная кора |
| 5 | Орбитофронтальная префронтальная кора | ||
| 6 | Вентральная префронтальная кора | 13 | Вентральная префронтальная кора |
| 7 | Предклинье | 14 | Предклинье |
| 8 | Поясная кора | 15 | Поясная кора |
| 9 | Медиальная задняя префронтальная кора | 16 | Медиальная задняя префронтальная кора |
Рис. 2.
Фронто-париетальная сеть контроля согласно атласу парцилляции коры головного мозга относительно 7 функциональных сетей, выделенных методом анализа функциональных связей в состоянии покоя на 1000 испытуемых [Yeo et al., 2011].
Fig. 2. The fronto-parietal control network, one of 7 networks obtained through functional connectivity analysis of resting-state fMRI data in 1000 subjects (cerebral cortex parcellation atlas by Yeo et al. [2011]).
Статистический анализ
Корреляционный анализ проводили между средневзвешенными показателями FA, RD, AD в 13 трактах (исключили кортикоспинальный тракт, нижний продольный пучок и затылочные щипцы) и 12 нейропсихологическими показателями. В качестве статистики использовался коэффициент корреляции Спирмана, полученные уровни значимости коэффициентов корреляции корректировались на множественные сравнения по количеству анализируемых трактов и нейропсихологических показателей по методу FDR (q = 0.05).
Сопоставление нейропсихологических данных с данными “ROI-to-ROI” анализа
С помощью общей линейной модели со случайными эффектами были получены связи между всеми возможными парами зон интереса фронто-париетальной сети на групповом уровне (для каждого полушария отдельно). Далее в модель были включены ковариаты, отражающие индивидуальные значения указанных выше нейропсихологических показателей. Для каждого нейропсихологического показателя строилась собственная линейная модель; вычислялись T-контрасты. К полученным уровням значимости применялась поправка на множественные сравнения по количеству всех анализируемых связей между парами зон интереса и количеству нейропсихологических показателей по методу FDR (q = 0.05).
РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЙ
Полученные нейропсихологические данные (средние по группе), с одной стороны, и результаты анализа корреляций между нейропсихологическими данными и особенностями трактов головного мозга и данными фМРТ представлены в табл. 2, в которую были включены все коэффициенты корреляции, чьи нескорректированные уровни значимости оказались меньше 5%.
Таблица 2.
Средние и стандартные отклонения нейропсихологических показателей, результаты корреляционного анализа между нейропсихологическими показателями и особенностями трактов головного мозга Table 2. Mean and standard deviations for neuropsychological indexes, the results of correlation analysis between neuropsychological indexes and characteristics of white matter tracts
| Субтест | Ср. | Стд. Откл. | МРТ данные | Статистика |
|---|---|---|---|---|
| Струп 1 (называние цветов), время | 27.0 | 3.7 | RD передней таламической лучистости левого полушария, мм2/сек | p = 0.0006, rs = 0.762 |
| FA передней таламической лучистости левого полушария | p = 0.004, rs = –0.671 | |||
| Струп 2 (чтение слов), время | 21.0 | 2.3 | ||
| Струп 3 (конфликтная проба), время | 41.8 | 10.2 | RD лобные щипцы, мм2/сек | p = 0.004, rs = 0.673 |
| Струп 4 (проба с переключением), время | 50.9 | 9.0 | ||
| Показатель “оттормаживание”, время | 14.8 | 7.5 | ||
| Показатель “переключение”, время | 9.2 | 9.4 | связанность височного и цингулярного компонентов фронто-париетальной сети правого полушария | p = 0.0001* |
| Визуальная беглость 1 – кол-во верных ответов | 12.3 | 3.5 | ||
| Визуальная беглость 2 – кол-во верных ответов | 13.3 | 4.3 | FA лобные щипцы | p = 0.003, rs = 0.687 |
| Визуальная беглость 3 – кол-во верных ответов | 10.1 | 5.0 | RD передней таламической лучистости левого полушария, мм2/сек | p = 0.004, rs = –0.672 |
| FA передней таламической лучистости левого полушария | p = 0.008, rs = 0.640 | |||
| RD лобные щипцы, мм2/сек | p = 0.005, rs = –0.669 | |||
| FA лобные щипцы | p = 0.004, rs = 0.675 | |||
| FA переднего поясного пучка левого полушария | p = 0.007, rs = 0.646 | |||
| Вербальная беглость 1 (фонетическая) – кол-во верных ответов | 54.5 | 9.9 | ||
| Вербальная беглость 2 (семантическая), кол-во верных ответов | 44.1 | 6.7 | ||
| Вербальная беглость 3 (семантическая с переключением), кол-во верных ответов | 6.2 | 1.9 |
Примечание: * – результат прошел коррекцию на множественность сравнений; FA – фракционная анизотропия, RD – радиальная диффузия, AD – аксиальная диффузия. Note: * – the result remained significant after correction for multiple comparisons; FA – fractional anisotropy, RD – radial diffusion, AD – axial diffusion.
Сопоставление нейропсихологических данных с данными диффузионно-взвешенной томографии не выявило результатов, прошедших коррекцию на множественность сравнений (q(FDR-corrected) = 0.05). Минимальные величины (р = 0.0006, проходит коррекцию q(FDR-corrected) = 0.1) были обнаружены для корреляции между радиальной диффузией левой передней таламический лучистости и показателем 1 субтеста теста Струпа (называние цветов).
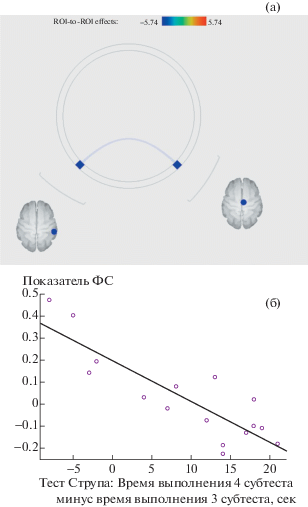
При анализе данных фМРТ покоя было обнаружено, что большие показатели разницы времени выполнения четвертого и третьего субтестов теста Струпа (слабость переключения) ассоциировались с меньшей связанностью между височным и поясным компонентами фронто-париетальной сети в правом полушарии (p-uncorrected = 0.0001, q(FDR-corrected) < 0.05) (рис. 3 а, б).
Рис. 3.
Результат “ROI-to-ROI” анализа: большие показатели разницы времени выполнения четвертого и третьего субтестов теста Струпа ассоциировались с меньшей связанностью между височным и поясным компонентами фронто-париетальной сети в правом полушарии (p-uncorrected = 0.0001, q(FDR-corrected) < 0.05) (а), диаграмма рассеяния с линией аппроксимации (линейная регрессия) – по оси абсцисс – “Время выполнения 4 субтеста Струпа минус время выполнения третьего субтеста”, по оси ординат – показатель функциональной связанности (б).
Fig. 3. Results of the “ROI-to-ROI” analysis: the more the difference in performance time between subtests 3 and 4 of the Stroop test, the less the functional connectivity between the temporal and cingulate components of the fronto-parietal network in the right hemisphere (p-uncorrected = = 0.0001) (а), scatterplot with trend line (linear regression): “Performance time of Stroop subtest 4 minus performance time of Stroop subtest 3” is on the X-axis, functional connectivity index is on the Y-axis (б).
Статистически значимых корреляций (даже на уровне номинальных) между показателями связанности компонентов фронто-париетальной сети и другими нейропсихологическими характеристиками выявлено не было.
ОБСУЖДЕНИЕ РЕЗУЛЬТАТОВ
Интерпретация низких показателей связанности между височным и поясным компонентами фронто-париетальной сети в правом полушарии при слабости переключения в тесте Струпа является дискуссионной.
Поясная извилина участвует в выявлении ситуаций конфликта, при которых необходим произвольный контроль; например, в классическом тесте Струпа активация передней поясной коры больше при ответе на конфликтные стимулы по сравнению с конгруэнтными [Van Veen, Carter, 2002; Gilbert, Burgess, 2008]. Несмотря на то что нейропсихологический показатель (разница выполнения субтестов 4 и 3 теста Струпа), корреляции с которым были выявлены, оценивает преимущественно переключение, зона, связанная с оттормаживанием, также могла оказаться задействованной потому, что оттормаживание в целом необходимо для успешного выполнения теста Струпа.
Участие височных отделов в переключении было также показано в литературе. Например, мета-анализ Kim et al. [2012] тридцати шести фМРТ-исследований с различными задачами на переключение показал, что переключение, независимо от конкретной задачи, было связано с активацией в задних нижних височных отделах (поле Бродмана 37). В маску фронто-париетальной сети Yeo и др. [2011], использованную в нашем исследовании, входят височные компоненты именно такой локализации.
Известно, что нижние височные отделы принимают участие в процессах зрительной рабочей памяти [Ranganath et al., 2004]. Можно предположить, что в нашем исследовании функциональная связанность поясного и височного отделов фронто-париетальной сети оказалась значимой для выполнения задачи на переключение, поскольку последние были вовлечены в удержание инструкции в зрительной модальности.
Неожиданным оказалось отсутствие корреляций (прошедших коррекцию на множественность сравнений) между связанностью дорсолатеральной префронтальной коры как элемента фронто-париетальной сети и тестируемыми характеристиками управляющих функций. Эти данные в какой-то степени противоречат результатам ранее проведенной работы, где было показано, что фактор оттормаживания (в этом исследовании это были данные факторного анализа большого блока различных нейропсихологических методик) коррелирует с функциональной связанностью между левой дорсолатеральной префронтальной корой и областью в постцентральной и незначительно прецентральной извилине слева, а также в дополнительной моторной области и передней поясной коре билатерально [Паникратова Я.Р. и др., 2017]. Возможной причиной является как использование в нашей работе других ROI, так и проведение анализа для отдельных, а не интегрированных показателей.
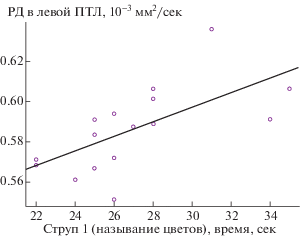
Хотя результаты корреляционного анализа между показателями трактов и управляющих функций не прошли коррекцию на множественность сравнений, авторы считают необходимым рассмотреть взаимосвязь радиальной диффузии в левой передней таламической лучистости (ПТЛ) и временем выполнения 1 серии теста Струпа (единственная корреляция, которая проходит коррекцию при использовании менее консервативного показателя q = 0.1; рис. 4). С одной стороны, по мнению ряда исследователей, в работах с использованием структурной, диффузионно-взвешенной и функциональной МРТ и с применением когнитивных тестов, и при большом количестве проверяемых гипотез, использование поправки FDR c q = 0.1 может быть обоснованным [van der Plas et al. 2017; Hellyer et al. 2015; Dyke et al. 2017], с другой – согласно проведенным нами исследованиям именно этот показатель является ключевым для процессов, ассоциируемых с высоким риском развития шизофрении [Томышев и др., 2016; Лебедева и др., 2019 в печати].
Рис. 4.
Диаграмма рассеяния с линией линейной регрессии – по оси ординат – “Радиальная диффузия в левой передней таламической лучистости (10–3 мм2/сек)”, по оси абсцисс – “Время называния в секундах в субтесте 1 теста Струпа” (rs – коэффициент корреляции Спирмана между показателями, указанными на диаграмме, p – уровень значимости данного коэффициента).
Fig. 4. A scatter plot with a linear regression line: “Radial diffusion in the left anterior thalamic radiation (10–3 mm2/s)” is on the Y-axis, “Naming time in seconds in the subtest 1 of the Stroop test” is on the X-axis (rs – is the Spearman correlation coefficient between the indicators on the scatterplot, p is the significance level of this coefficient).
Известно, что показатель радиальной диффузии связывается с состоянием миелиновых оболочек трактов белого вещества [Aung et al., 2013]. Можно предположить, что скорость называния цветов коррелировала с вариабельностью этой характеристики у здоровых испытуемых в левой ПЛТ – тракте, который соединяет медио-дорсальные и передние ядра таламуса с префронтальной корой [Coenen et al., 2012]. В целом фронто-таламическая система рассматривается как одна из важнейших управляющих систем мозга. Ее работу связывают с избирательной настройкой корковых нейронных сетей на обработку значимой информации [Р.И. Мачинская, 2015]. Это может объяснять корреляцию показателей состояния тракта с называнием цветов в тесте Струпа, представляющим собой простую, но все же произвольную деятельность, требующую избирательного внимания [Barch, et al., 2004].
ВЫВОДЫ
В рамках настоящего исследования выявлены единичные корреляции, в целом, полученные результаты согласуются с существующими представлениями о роли фронто-таламических трактов в обеспечении скорости обработки информации, а также о значимости функциональной связанности поясной извилины и височных отделов при выполнении задач с нагрузкой на зрительное избирательное внимание.
Исследование было поддержано грантом Российского фонда фундаментальных исследований № 15-06-05758.
Список литературы
Лурия А.Р. Основы нейропсихологии. М.: Изд-во МГУ, 1973. 374 с.
Лурия А.Р. Высшие корковые функции и их нарушение при локальных поражениях мозга. М.: Изд-во МГУ, 1962. 431 с.
Мачинская Р.И. Управляющие системы мозга. Журнал высшей нервной деятельности им. И.П. Павлова. 2015. 65 (1): 33–61.
Мачинская Р.И. Формирование нейрофизиологических механизмов произвольного избирательного внимания у детей младшего школьного возраста. Москва. 2001. 278 с.
Паникратова Я.Р., Власова Р.М., Синицын В.Е., Печенкова Е.В. Состояние регуляторных функций и функциональные связи дорсолатеральной префронтальной коры. Материалы конференции “Когнитивная наука в Москве: новые исследования”. БукиВеди, ИППиП. 2017. 266–271.
Томышев А.С., Лебедева И.С., Ахадов Т.А., Омельченко М.А., Ублинский М.В., Семенова Н.А., Каледа В.Г. МРТ-исследование особенностей проводящих путей головного мозга у пациентов c ультравысоким риском развития эндогенных психозов. Бюллетень экспериментальной биологии и медицины. 2016. 162 (10): 417–421.
Alvarez Julie A., Emory Eugene. Executive Function and the Frontal Lobes: A Meta-Analytic Review. Neuropsychology Review. 2006. 16 (1): 17–42.
Aung W.Y., Mar S., Benzinger T.L. Diffusion tensor MRI as a biomarker in axonal and myelin damage. Imaging Med. 2013. 5 (5): 427–440.
Barch D.M., Carter C.S., Cohen J.D. Factors influencing Stroop performance in schizophrenia. Neuropsychology. 2004. 18 (3): 477–84.
Behrens T.E., Berg H.J., Jbabdi S., Rushworth M.F., Woolrich M.W. Probabilistic diffusion tractography with multiple fibre orientations: What can we gain? Neuroimage. 2007. 34 (1): 144–155.
Bellebaum C., Daum I. Cerebellar involvement in executive control. The Cerebellum. 2007. 6 (3): 184–192.
Chen T., Becker B., Camilleri J., Wang L., Yu S., Eickhoff S.B., Feng C. A domain-general brain network underlying emotional and cognitive interference processing: evidence from coordinate-based and functional connectivity meta-analyses. Brain Structure and Function. 2018. 223 (8): 3813–3840.
Chung H.J., Weyandt L.L., Swentosky A. The physiology of executive functioning. Handbook of executive functioning. Springer New York, 2014. 13–27.
Coenen V.A., Schlaepfer T.E., Allert N., Mädler B. Diffusion tensor imaging and neuromodulation: DTI as key technology for deep brain stimulation. Int Rev Neurobiol. 2012. 107: 207–234.
Collette F., Van der Linden M., Laureys S., Delfiore G., Degueldre C., Luxen A., Salmon E. Exploring the unity and diversity of the neural substrates of executive functioning. Human Brain Mapping. 2005. 25 (4): 409–423.
Delis D.C., Kramer J.H., Kaplan E., Holdnack J. Reliability and validity of the Delis-Kaplan Executive Function System: an update. J IntNeuropsychol Soc. 2004. 10 (2): 301–3.
Dyke J.P., Cameron E., Hernandez N., Dydak U., Louis E.D. Gray matter density loss in essential tremor: a lobule by lobule analysis of the cerebellum. Cerebellum Ataxias. 2017. 3: 4–10.
Friedman N.P., Miyake A. Unity and diversity of executive functions: Individual differences as a window on cognitive structure. Cortex. 2017. 86: 186–204.
Garavan H., Ross T.J., Murphy K., Roche R.A.P., Stein E.A. Dissociable executive functions in the dynamic control of behavior: inhibition, error detection, and correction. Neuroimage. 2002. 17 (4): 1820–1829.
Gilbert S.J., Burgess P.W. Executive function. Curr Biol. 2008. 18 (3): 110–114.
Grieve S.M., Williams L.M., Paul R.H., Clark C.R., Gordon E. Cognitive aging, executive function, and fractional anisotropy: a diffusion tensor MR imaging study. AJNR Am J Neuroradiol. 2007. 28 (2): 226–35.
Hellyer P.J., Scott G., Shanahan M., Sharp D.J., Leech R. Cognitive Flexibility through Metastable Neural Dynamics Is Disrupted by Damage to the Structural Connectome. J Neurosci. 2015. 35 (24): 9050–9063.
Kim C., Cilles S.E., Johnson N.F., Gold B.T. Domain general and domain preferential brain regions associated with different types of task switching: a meta-analysis. Hum Brain Mapp. 2012. 33 (1): 130–42.
Lebel C., Warner T., Colby J., Soderberg L., Roussotte F., Behnke M., Davis Eyler F., Sowell E.R. White matter microstructure abnormalities and executive function in adolescents with prenatal cocaine exposure. Psychiatry Res. 2013. 213 (2): 161–168.
Mazoyer B., Zago L., Mellet E., Bricogne S., Etard O., Houdé O., Crivello F., Joliot M., Petit L., Tzourio-Mazoyer N. Cortical networks for working memory and executive functions sustain the conscious resting state in man. Brain research bulletin. 2001. 54 (3): 287–298.
Mesulam M.M. Large-scale neurocognitive networks and distributed processing for attention, language, and memory. Ann Neurol. 1990. 28 (5): 597–613.
Monchi O., Petrides M., Strafella A.P., Worsley K.J., Doyon J. Functional role of the basal ganglia in the planning and execution of actions. Annals of Neurology: Official Journal of the American Neurological Association and the Child Neurology Society. 2006. 59 (2): 257–264.
Niendam T.A., Laird A.R., Ray K.L., Dean Y.M., Glahn D.C., Carter C.S. Meta-analytic evidence for a superordinate cognitive control network subserving diverse executive functions. Cognitive, Affective, & Behavioral Neuroscience. 2012. 12 (2): 241–268.
Nowrangi M.A., Lyketsos C., Rao V., Munro C.A. Systematic review of neuroimaging correlates of executive functioning: converging evidence from different clinical populations. J Neuropsychiatry ClinNeurosci. 2014. 26 (2): 114–125.
Park H.J., Friston K. Structural and functional brain networks: from connections to cognition. Science. 2013. 342 (6158): 579–589.
Petersen S.E., Posner M.I. The attention system of the human brain: 20 years after. Annual review of neuroscience. 2012. 35: 73.
Ptak R., Di Pietro M., Schnider A. The neural correlates of object-centered processing in reading: a lesion study of neglect dyslexia. Neuropsychologia. 2012. 50 (6): 1142–1150.
Ranganath C., Cohen M.X., Dam C., D’Esposito M. Inferior temporal, prefrontal, and hippocampal contributions to visual working memory maintenance and associative memory retrieval. J Neurosci. 2004. 24 (16): 3917–3925.
Reineberg A.E., Banich M.T. Functional connectivity at rest is sensitive to individual differences in executive function: A network analysis. Human brain mapping. 2016. 37 (8): 2959–2975.
Sasson E., Doniger G.M., Pasternak O., Tarrasch R., Assaf Y. White matter correlates of cognitive domains in normal aging with diffusion tensor imaging. Front Neurosci. 2013. 7 (32): 1–13.
Seeley W.W., Menon V., Schatzberg A.F., Keller J., Glover G.H., Kenna H., Greicius M.D. Dissociable intrinsic connectivity networks for salience processing and executive control. Journal of Neuroscience. 2007. 27 (9): 2349–2356.
Seghete K.L.M., Hertinga M.M., Nagel B.J. White matter microstructure correlates of inhibition and task-switching in adolescents. Brain Research. 2013. 1527: 15–28.
Smolker H.R., Depue B.E., Reineberg A.E., Orr J.M., Banich M.T. Individual differences in regional prefrontal gray matter morphometry and fractional anisotropy are associated with different constructs of executive function. Brain Struct Funct. 2015. 220 (3): 1291–306.
Stuss D.T. Functions of the frontal lobes: relation to executive functions //Journal of the international neuropsychological Society. 2011. 17 (5): 759–765.
Sylvester C.Y.C., Wager T.D., Lacey S.C., Hernandez L., Nichols T.E., Smith E.E., Jonides J. Switching attention and resolving interference: fMRI measures of executive functions. Neuropsychologia. 2003. 41 (3): 357–370.
Szameitat A.J., Schubert T., Müller K., Von Cramon D.Y. Localization of executive functions in dual-task performance with fMRI. Cognitive Neuroscience, Journal of. 2002. 14 (8): 1184–1199.
Takeuchi H., Taki Y., Sassa Y., Hashizume H., Sekiguchi A., Fukushima A., Kawashima R. Verbal working memory performance correlates with regional white matter structures in the frontoparietal regions. Neuropsychologia. 2011. 49 (12): 3466–3473.
Takeuchi H., Taki Y., Nouchi R., Sekiguchi A., Hashizume H., Sassa Y., Kotozaki Y., Miyauchi C.M., Yokoyama R., Iizuka K., Nakagawa S., Nagase T., Kunitoki K., Kawashima R. Degree centrality and fractional amplitude of low-frequency oscillations associated with Stroop interference. Neuroimage. 2015. 119: 197–209.
van der Plas E., Schachar R.J., Hitzler J., Crosbie J., Guger S.L., Spiegler B.J., Ito S., Nieman B.J. Brain structure, working memory and response inhibition in childhood leukemia survivors. Brain Behav. 2016. 7 (2): 1–16.
Van Veen V., Carter C. S. The anterior cingulate as a conflict monitor: fMRI and ERP studies. Physiology & behavior. 2002. 77 (4): 477–482.
Vollmer B., Lundequist A., Martensson G., Nagy Z., Lagercrantz H., Smedler A.C., Forssberg H. Correlation between white matter microstructure and executive functions suggests early developmental influence on long fibre tracts in preterm born adolescents. PLoS One. 2017. 12 (6): 1–16.
Whitfield-Gabrieli S., Nieto-Castanon A. Conn: a functional connectivity toolbox for correlated and anticorrelated brain networks. Brain Connect. 2012. 2 (3): 125–141.
Yendiki A., Panneck P., Srinivasan P., Stevens A., Zollei L., Augustinack J. Automated probabilistic reconstruction of white-matter pathways in health and disease using an atlas of the underlying anatomy. Frontiers in Neuroinformatics, 2011. 5 (23).
Yeo B.T., Krienen F.M., Sepulcre J., Sabuncu M.R., Lashkari D., Hollinshead M., Roffman J.L., Smoller J.W., Zöllei L., Polimeni J.R., Fischl B., Liu H., Buckner R.L. The organization of the human cerebral cortex estimated by intrinsic functional connectivity. J Neurophysiol. 2011. 106 (3): 1125–1165.
Zanto T.P., Gazzaley A. Fronto-parietal network: flexible hub of cognitive control. Trends Cogn Sci. 2013. 17 (12): 602–3.
Zeki S., Shipp S. The functional logic of cortical connections. Nature. 1988. 335 (6188): 311–317.
Дополнительные материалы отсутствуют.
Инструменты
Журнал высшей нервной деятельности им. И.П. Павлова