

Журнал высшей нервной деятельности им. И.П. Павлова, 2019, T. 69, № 1, стр. 65-77
ИССЛЕДОВАНИЕ КОГНИТИВНЫХ СПОСОБНОСТЕЙ У МАКАК-РЕЗУСОВ
Д. Н. Подвигина 1, 2, *, Л. Е. Иванова 1, И. А. Варовин 1, А. К. Хараузов 1
1 Институт физиологии им. И.П. Павлова РАН
Санкт-Петербург, Россия
2 Санкт-Петербургский государственный университет
Санкт-Петербург, Россия
* E-mail: daria-da@yandex.ru
Поступила в редакцию 27.02.2017
После доработки 18.01.2018
Принята к публикации 05.03.2018
Аннотация
В поведенческих экспериментах исследовали способность обезьян Macaca mulatta к выполнению когнитивных тестов разной степени сложности, предъявляемых на сенсорном экране компьютера. Задача состояла в распознавании доминирующей ориентации в текстурах различной степени упорядоченности. Анализировали скорость обучения, процент правильных ответов и время решения задачи в зависимости от ее сложности. Параллельно у животных оценивали выраженность исследовательской активности по реакции на незнакомые предметы (тест “реакция на новый объект”). Процент правильных ответов увеличивался с упорядоченностью тестовых изображений и был сопоставим с результатами, полученными ранее на человеке в сходных экспериментальных условиях. У обезьян индекс исследовательской активности коррелировал со скоростью обучения и был ниже у тех, кто склонен к отвлечению внимания, то есть с большим временем ответа. Полученные на обезьянах результаты позволяют моделировать нейронные процессы, протекающие в головном мозге человека при распознавании образов, от кодирования физических свойств изображений до принятия решений. Взаимосвязь между уровнем исследовательской активности и скоростью обучения позволяет использовать тест “реакция на новый объект” при отборе перспективных особей для участия в поведенческих экспериментах, что существенно сократит время, затраченное на обучение обезьян новым задачам.
Понимание нейрофизиологических основ функционирования мозга человека невозможно без использования животных моделей с применением инвазивных методов исследования. Наиболее близкими животными к человеку в эволюционном плане, а также по анатомическому строению и физиологическим особенностям, являются обезьяны. Среди всех известных видов обезьян макаки-резусы представляют собой наиболее распространенную животную модель для изучения нейронных процессов [Freeman, Gosling, 2010]. Однако, учитывая явные различия во многих когнитивных способностях человека и обезьяны, в размере и строении их мозга, мы не всегда можем напрямую обобщать получаемые результаты [Passingham, 2009]. Поэтому при изучении различных функций мозга животных, перед тем как интерполировать данные на другие виды, в том числе на человека, мы должны убедиться, что исследуемые процессы протекают сходным образом, например, путем сопоставления результативности выполнения экспериментальных тестов.
На примере функционирования зрительной системы человека и обезьяны были проведены многочисленные сравнения результативности работы первичных звеньев зрительного анализатора, осуществляющих описание физических характеристик изображений. В частности, было показано сходство кривых контрастной чувствительности, одинаковые способности к различению цвета, ориентации и размера фигур [De Valois et al., 1974a; 1974b; Vázquez et al., 2000; Schmitt et al., 2013]. Сравнение функций высших отделов зрительного анализатора, таких как инвариантное распознавание изображений, способность выделять объекты из окружения, привело к аналогичным выводам о том, что в основе этих процессов у человека и обезьяны лежат одинаковые нейронные механизмы [Rajalingham et al., 2015]. Успешность решения когнитивных тестов, направленных на определение пространственного и количественного соотношения предметов и понимание несложных причинно-следственных связей, одинакова у разных видов приматов, включая человека [Hermann et al., 2007]. Однако оказалось, что перенос обучения в задачах зрительного различения у обезьян, в отличие от человека, зависит от типа информации, содержащегося в зрительных стимулах. Так, обезьяны, обученные решению задач М. Бонгарда [Бонгард, 1967], испытывали сложности с переносом результатов обучения при работе с группой стимулов, различающихся по пространственным отношениям между элементами изображений [Дудкин, 2007].
Задачей нашего исследования было продолжение изучения когнитивных способностей обезьян и сопоставление результатов с данными, полученными на человеке в сходных экспериментальных условиях. В качестве тестового материала использовали набор изображений – текстур, различающихся по степени ориентационной упорядоченности составляющих элементов, где задачей испытуемых было определение доминирующей ориентации в стимуле. Преимущество такого набора стимулов в том, что изменяя упорядоченность текстур, можно контролировать сложность задачи, не меняя при этом основные физические характеристики изображений. Ранее с помощью этих тестов были исследованы зависимости психофизических и электрофизиологических показателей от сложности задачи у человека [Хараузов и др., 2007; Сельченкова и др., 2011; Harauzov et al., 2016], локализованы области мозга, активирующиеся при работе с данными изображениями [Шелепин и др., 2009]. Использование аналогичных экспериментальных парадигм у обезьян и человека существенно расширит наши возможности по изучению нейрофизиологических механизмов решения сложных пространственных задач.
Другой целью нашего исследования было сопоставление индивидуальных особенностей поведения приматов со скоростью об-учения когнитивным задачам. Еще И.П. Павлов и его сотрудники, изучая условно-рефлекторные реакции собак, обращали внимание на индивидуальные различия в их поведении. Эти индивидуальные особенности условно-рефлекторной деятельности не могут быть объяснены исключительно разнообразием экспериментальных ситуаций, в их основе лежит сочетание фундаментальных свойств нервной системы, которое И.П. Павлов назвал типом нервной системы [Стреляу, 1982]. У обезьян индивидуальные различия также начали изучать в первой половине ХХ века [Crawford, 1938; Yerkes, 1939; Hebb, 1946] на основании наблюдений и результатов поведенческих экспериментов. В этих и других, более поздних работах анализируются индивидуальные различия в поведении животных, характеризующиеся постоянством, не зависящие от ситуации и определяющие то, что можно назвать индивидуальностью [Réale et al., 2007; Griffin et al., 2015] или темпераментом [Heath-Lange et al., 1999; Coleman et al., 2005].
Не так давно исследователи затронули вопрос о взаимосвязи когнитивных способностей обезьян и индивидуальных особенностей поведения. Так, в работе Coleman и соавт. была показана зависимость скорости обучения макак несложным манипуляциям с физическими объектами от их темперамента, а точнее, от особенностей исследовательского поведения, которые оценивали по реакции на незнакомый предмет [Coleman et al., 2005]. В нашей работе мы развили предложенный метод оценки исследовательской реакции, что позволило более точно определять выраженность исследовательской активности у обезьян и сопоставлять со способностью к обучению работе в виртуальном пространстве с изображениями, предъявляемыми на сенсорном мониторе компьютера.
МЕТОДИКА
Исследования проводили на пяти самцах Macaca mulatta в возрасте 6–9 лет. Все животные были рождены и выращены в неволе и имели опыт участия в поведенческих экспериментах. Протоколы опытов были утверждены Комиссией по гуманному обращению с животными Института физиологии им. И.П. Павлова РАН. В описании результатов обезьяны обозначены первой буквой своей клички и возрастом на момент проведения экспериментов: Ф6, Л8, Н8, Ю9 и Ф9.
Обучение и проведение когнитивных тестов осуществляли с помощью экспериментального комплекса для проведения психофизических и поведенческих исследований на обезьянах, разработанного в Институте физиологии им. И.П. Павлова РАН. Основу комплекса составляет компьютер с сенсорным дисплеем, что позволяет проводить зрительную и слуховую стимуляцию любой сложности, автоматически регистрировать ответную реакцию и обеспечивать пищевое и питьевое подкрепление в случае правильного ответа. Во время эксперимента обезьяна находится в специальной передвижной клетке из прозрачного оргстекла с отверстием для передних конечностей и встроенными системами подкрепления. Расстояние между клеткой и сенсорным экраном составляет 20 см. Более подробно экспериментальная установка описана в статье Ивановой и соавт. [Иванова и др., 2016].
Когнитивные способности обезьян изучали с помощью набора тестов, разработанных в Институте физиологии им. И.П. Павлова РАН для электрофизиологических и томографических исследований на человеке в задачах оценки статистических свойств сложных изображений [Хараузов и др., 2007; Шелепин и др., 2009; Harauzov et al., 2016]. Тесты представляют собой набор матриц, каждая из которых состоит из 100 элементов – решеток Габора разной ориентации. Матрицы различаются по количеству однонаправленных элементов, другими словами, по степени упорядоченности (рис. 1). Всего в наборе имеется восемь типов матриц, содержащих 100, 64, 56, 48, 40, 32, 24 и 16 однонаправленных элементов. Каждый тип матриц (кроме полностью упорядоченных) имеет по 40 разных реализаций, различающихся взаимным расположением элементов, но имеющих одинаковую степень упорядоченности. Доминирующая ориентация в матрицах может быть либо вертикальной, либо горизонтальной. Задача испытуемых – определить, каких элементов в изображении больше: вертикальных или горизонтальных. Как видно из рис. 1, по мере уменьшения количества однонаправленных элементов задача категоризации матрицы и отнесения ее к “вертикальному” или “горизонтальному” типу усложняется. Преимущество данного набора стимулов в том, что изменение сложности можно легко контролировать, при этом основные физические характеристики стимулов, такие как яркость, контраст, размер и так далее, остаются постоянными.
Рис. 1.
Примеры тестовых стимулов – матриц различной степени упорядоченности. Цифры у каждой матрицы обозначают относительное количество однонаправленных элементов. Задача – определить доминирующую ориентацию.
Fig. 1. The examples of test stimuli – matrixes with different complexity level. The numbers are the percentage of the co-oriented elements within one matrix. The task was to identify the dominant orientation.

Обучение обезьян выполнению когнитивных тестов проходило в несколько этапов. На первом этапе в левой и в правой части экрана предъявляли две черно-белые решетки, одна из которых имела вертикальную, другая – горизонтальную ориентацию. Методом положительного подкрепления животных обучали выбирать вертикальную решетку. В зависимости от местоположения целевого стимула (вертикальной решетки) на сенсорном экране выбирали области, прикосновение к которым фиксируется как правильный ответ. В случае правильного ответа автоматически выдавалось пищевое подкрепление. Нажатие на любую другую область экрана не подкреплялось. Сразу после касания экрана стимулы исчезали, и через 5 с на экране появлялась новая пара решеток. Взаимное расположение вертикальной и горизонтальной решеток в каждом предъявлении было случайным. Тренировки проводили по одному разу в каждый рабочий день, по 30 мин для каждой обезьяны.
После достижения уровня правильных ответов 75% проводили еще 2–4 тренировки с теми же стимулами, затем решетки заменяли максимально упорядоченными матрицами, состоящими исключительно из горизонтальных или вертикальных элементов Габора. Угловые размеры матриц составляли 27 угловых градусов, расстояние между двумя матрицами – 10 угловых градусов. Как и на этапе с решетками, подкрепляли выбор только вертикальных матриц. В паре с целевым стимулом – вертикальной матрицей – предъявляли матрицу, состоящую из горизонтальных элементов. После того, как обезьяны обучались работе с максимально упорядоченными изображениями, в которых все 100 элементов Габора имели одинаковую ориентацию, им начинали предъявлять матрицы, содержащие 64 элемента с одинаковой ориентацией. Затем, по мере обучения, количество однонаправленных элементов в матрицах снижали до 48, затем до 40 и до 32. Во всех случаях критерием того, что обезьяна усвоила данную задачу, считали достижение уровня 75% правильных ответов [Альтман, 1990], после которого проводили еще несколько тренировочных сессий для закрепления результата и переходили на более сложные стимулы.
После обучения обезьян работе с матрицами одинаковой степени упорядоченности переходили к тренировке для выполнения основных экспериментов, в которых использовали сразу четыре набора матриц разной степени упорядоченности, содержащих 40, 32, 24 и 16 однонаправленных элементов. Как и в процессе обучения, обезьяне предъявляли две матрицы с одинаковой степенью упорядоченности, но с разной доминирующей ориентацией в левой и в правой половине экрана. После фиксации ответной реакции предъявляли следующую пару матриц с другим количеством однонаправленных элементов. Взаимное расположение “вертикальных” и “горизонтальных” матриц на экране и последовательность предъявления пар матриц с определенной степенью упорядоченности менялись случайным образом. Для получения стабильного уровня правильных ответов с каждой обезьяной провели по 20 тренировочных сессий в режиме предъявления матриц разной степени упорядоченности, после чего перешли к основной экспериментальной части исследования.
Всего на каждой макаке был проведен 31 эксперимент, по одному в день, длительностью по 30 мин. В каждом эксперименте обезьяны успевали сделать в среднем по 100 нажатий на экран. По результатам каждого эксперимента анализировали процент правильных ответов и время ответа. Мы используем термин “время ответа” вместо “времени реакции”, поскольку термин “время реакции” подразумевает необходимость давать ответ как можно скорее. В нашем случае обезьяны не были ограничены во времени и могли нажать на экран через минуту после предъявления стимула.
Для выявления индивидуальных особенностей в поведении обезьян был взят за основу метод оценки исследовательского инстинкта, предложенный Coleman и соавт. [Coleman et al., 2005]. Суть метода заключалась в анализе времени реакции на новый, незнакомый для обезьяны предмет. В наших исследованиях количество незнакомых предметов было увеличено до пяти. В основном это были разноцветные детские игрушки. Каждый предмет по одному ставили перед обезьяной и фиксировали время до первого касания. Сразу после того, как обезьяна дотронулась до предмета, его заменяли другим, и так далее. Если обезьяна игнорировала какой-либо из предметов и не брала его в течение трех минут, фиксировали отсутствие реакции на него и заменяли другим предметом. Во время эксперимента обезьяна находилась в прозрачной экспериментальной камере с широким отверстием для передних конечностей. Камера и окружающая обстановка были знакомы обезьянам, эксперимент проводили два знакомых для них исследователя, находящихся в одном метре от клетки. Реакцию на предъявление каждого из пяти предметов записывали на видеокамеру и на основе видеозаписи оценивали время до касания.
Обработку и анализ результатов проводили с использованием пакета программ Microsoft Excel 2013. Для проверки статистической достоверности результатов использовали двусторонний t-критерий Стьюдента и коэффициент корреляции Пирсона.
РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЯ
В процессе обучения обезьян определению доминирующей ориентации в тестовых изображениях было обнаружено, что не все животные одинаково быстро справляются с задачами. Некоторые обезьяны успевали понять суть задачи за одну тренировочную сессию, у других обучение занимало несколько дней. На рис. 2 (а) приведены примеры разной скорости обучения у двух обезьян на матрицах, содержащих 64 однонаправленных элемента, предъявленных за одну тренировочную сессию. Каждый ответ принимали за +1, если он был правильный, и за –1, если ответ был ошибочный. Значения каждого ответа суммировали с предыдущими, начиная с нуля, в результате получили зависимость количества правильных ответов от порядкового номера предъявления. Если кривая колеблется около нуля, это означает, что животное делает ошибки в половине случаев. Отклонение в положительную сторону означает, что преобладают правильные ответы, и наоборот, отклонение в отрицательную сторону означает, что преобладают ошибочные ответы.
Рис. 2.
Скорость обучения обезьян выполнению когнитивных тестов. (а) – иллюстрация разной скорости обучения у двух обезьян (Ю9 и Л8) за одну тренировочную сессию. Цифры в скобках указывают процент правильных ответов, полученный в конце тренировки; (б) – среднее количество дней, потребовавшихся каждой обезьяне для обучения работе со всеми новыми стимулами.
Fig. 2. The rate of macaque learning to perform cognitive tests. (a) – an illustration of different learning rate of the two monkeys (Ю9 and Л8) during one training session. The numbers in brackets are the percentage of correct answers at the end of the training session; (б) – an average number of days each monkey needed to learn how to work with a new stimulus.
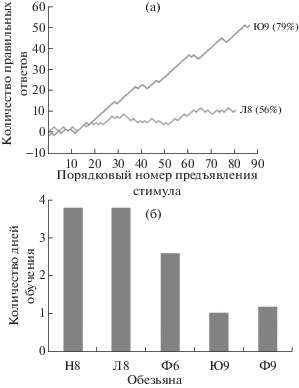
На примере видно, что животное Ю9 в ответ на первые 15 предъявлений совершает случайное гадание. Затем, после 15 предъявлений, начинает понимать суть задачи и далее отвечает в основном правильно, достигая к концу эксперимента 79% правильных ответов. Животное Л8 совершает гораздо больше ошибочных нажатий. Небольшое улучшение наблюдается после 25 предъявлений, однако в среднем количество правильных ответов составило у него в этот день 56%. Таким образом, несмотря на то, что в матрицах 64 можно без труда определить доминирующую ориентацию, для обезьяны Л8 переход с матрицы 100 на матрицу 64 вызвал затруднение. В результате делается вывод, что животное Ю9 обучилось работе с матрицей 64 за один день, а животному Л8 предстоит еще повторить обучение.
Обучение работе с каждой матрицей считалось успешным, если был достигнут семидесятипятипроцентный уровень правильных ответов. Мы подсчитали, сколько дней было необходимо каждой особи для обучения работе с каждым типом стимулов.
Наиболее успешными были обезьяны Ю9 и Ф9, им было достаточно одного-двух дней для обучения каждому типу матриц. Животному Ф9 потребовалось девять дней для перехода от решеток к матрицам 100, но затем дальнейшее усложнение задачи не вызывало у него трудностей: обучение переходу к менее упорядоченным матрицам происходило за одну тренировочную сессию. Самыми отстающими оказались обезьяны Н8 и Л8, им потребовалось в среднем четыре дня для обучения работе с каждым типом матриц, за исключением матрицы 40, которую они освоили за одну тренировку.
Рис. 2 (б) объединяет данные по числу дней, потребовавшихся каждой макаке для достижения 75% правильных ответов при работе со всеми типами матриц. На рисунке наглядно представлены различия в скорости обучения новым стимулам у разных макак.
Несмотря на разную скорость обучения, к концу проведения тренировок все пять макак успешно решали задачу определения доминирующей ориентации в матрицах, содержащих 40, 32, 24 и 16 однонаправленных элементов, которые предъявляли в случайном порядке во время одного эксперимента. Исключение составляли лишь самые сложные для распознавания матрицы (16), в ответ на которые только одна обезьяна превысила установленный нами порог обучения в 75%. У остальных макак количество правильных ответов на эти матрицы варьировало от 63% до 70% и не увеличивалось по мере тренировочного процесса. Таким образом, для выбранного вида животных матрицы, содержащие 16 однонаправленных элементов, являются стимулом, близким к порогу распознавания доминирующей ориентации.
После того, как в процессе тренировок все обезьяны стали показывать стабильные результаты, был проведен 31 эксперимент с матрицами 40, 32, 24 и 16 в условиях, аналогичных тренировочному процессу. Анализировали количество правильных ответов и время между предъявлением стимула и ответной реакцией у каждой обезьяны на каждую матрицу. На рис. 3 (а) представлены усредненные за все дни эксперимента данные по числу правильных ответов на каждый тип матриц.
Рис. 3.
Зависимость между результатами выполнения задачи на определение доминирующей ориентации в матрице и степенью ее упорядоченности. (а) – зависимость количества правильных ответов от степени упорядоченности матриц. По оси абсцисс – количество однонаправленных элементов в матрице, величина, обратно пропорциональная сложности задачи. По оси ординат – процент правильных ответов в условиях двухальтернативного выбора, 50% – уровень случайного гадания. Представлены данные по пяти обезьянам (Ф6, Л8, Н8, Ю9 и Ф9), усредненные на основе 31 измерения. Планки погрешности – ошибка среднего; (б) – зависимость времени реакции от степени упорядоченности матриц.
Fig. 3. The dependence between the results of performing the task on matrix dominant orientation detection and the matrix complexity. (a) – the dependence between the number of correct answers and the stimulus complexity level. X-axis – the number of co-oriented elements within a matrix, a value inversely proportional to the task complexity. Y-axis – the percentage of the correct answers, 50% – chance guessing level. The data of five monkeys (Ф6, Л8, Н8, Ю9, Ф9) averaged over 31 observations. Error bars are the error of mean; (б) – the dependence between the reaction time and the stimulus complexity level.
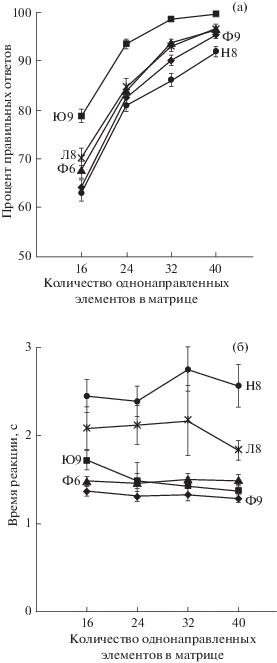
Как видно на рис. 3 (а), у всех обезьян процент правильных ответов увеличивается по мере увеличения степени упорядоченности стимула, то есть по мере упрощения задачи. В среднем результативность увеличивается с 69% в ответ на самые сложные матрицы до 96% в ответ на самые легкие для распознавания стимулы. На рисунке также наблюдаются различия в результативности между разными животными. В среднем по всем типам матриц наибольшее число правильных ответов (93%) показала обезьяна Ю9. Причем это единственная обезьяна, результативность которой выходит на плато по мере увеличения упорядоченности стимула: процент правильных ответов на матрицы 32 и 40 значимо не различается (98.6% и 99.6% соответственно, p = = 0.06, двусторонний Т-критерий, n = 31). Остальные четыре макаки справлялись с задачами в среднем на уровне 81–86%, значимо улучшая результативность ответов при каждом увеличении упорядоченности матриц (p < 0.05, двусторонний Т-критерий, n = 31).
При анализе времени ответа в качестве меры центральной тенденции использовали медиану. Поскольку животные могли отвлекаться и не всегда нажимали на экран сразу после предъявления стимула, распределение времени ответа было далеко от нормального (рис. 3 (б)). Поэтому медиана в данном случае подходит лучше среднего арифметического, так как нивелирует влияние больших задержек в ответах.
Для того чтобы избавиться от значительных выбросов в значениях времени ответа, возникающих при отвлечении обезьян от работы, для каждого животного в каждом эксперименте рассчитывали верхнюю внешнюю границу выборки как сумму третьего квартиля и межквартильного размаха, умноженного на 3 [NIST/SEMATECH e-Handbook of Statistical Methods]. Значения, превышающие верхнюю внешнюю границу, исключали из дальнейшего анализа. В среднем по всем обезьянам за все время проведения экспериментов таким образом было отброшено 2% отклоняющихся значений времени ответа.
На рис. 3 (б) представлены усредненные за все дни эксперимента данные по времени ответа на каждый тип матриц у каждой макаки. Значимая зависимость времени ответа от сложности задачи была обнаружена только у одного животного Ю9: чем сложнее задача, тем больше у него было время реакции (p < < 0.05, двусторонний Т-критерий, n = 31). Следует отметить, что именно это животное показало наилучшие результаты по количеству правильных ответов (см. рис. 3 (а)).
Животные Ю9, Ф9 и Ф6 имели близкие значения времени ответа, в среднем 1.4 с, в то время как животные Л8 и Н8 нажимали на экран значительно позже (2 и 2.5 с соответственно). Причем, как видно на рис. 3 (б), у обезьян Л8 и Н8, наряду с увеличением времени ответа, наблюдается также увеличение разбросов значений, полученных в разные дни эксперимента. Поэтому на основе приведенных графиков невозможно объяснить, чем вызвано увеличение времени ответа у двух животных. Это может быть либо следствием того, что для обезьян Л8 и Н8 задача слишком сложная и требует большего времени для принятия решения, либо из-за того, что они чаще отвлекаются.
Для того чтобы ответить на поставленный вопрос, мы проанализировали гистограммы распределения времени ответа в ответ на все типы матриц, регистрируемые в каждом отдельном эксперименте. В результате для каждой обезьяны было обработано более 3000 ответов, полученных за все дни наблюдения. Величина временных интервалов для построения гистограмм составила 1 с, за исключением самых коротких времен ответа, где интервал был 0.5 с. На рис. 4 представлены итоговые гистограммы, отсортированные по ширине распределения. Вверху представлены животные с наиболее стабильными временами ответов и соответственно с более узким распределением, внизу – обезьяны с большим числом отклонений во времени ответа соответственно с более широким распределением.
Рис. 4.
Гистограммы распределения времени реакции на все типы матриц за все время проведения экспериментов.
Fig. 4. The distributions of reaction time in responses to all matrix types over all experimental sessions.
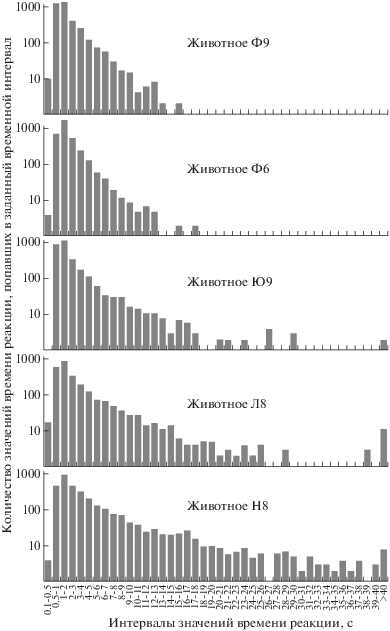
На рис. 4 видно, что наиболее широкое распределение значений наблюдается у обезьяны Н8, которая в среднем за все дни проведения экспериментов показала самое длительное время ответа. Вторая по ширине гистограмма принадлежит обезьяне Л8, также показавшей большое время ответа. Промежуточное значение занимает обезьяна Ю9, самые узкие распределения наблюдаются у макак Ф6 и Ф9. Следует отметить, что, несмотря на существенные различия в ширине гистограмм, максимум значений времени ответа у всех животных попадает в интервал 1–2 с. Эти наблюдения позволяют утверждать, что время от начала предъявления стимула до момента принятия решения и последующего развития моторной реакции у обезьян в предложенных экспериментальных условиях примерно одинаковое. Различия во времени ответа, выявленные на рис. 4, объясняются различиями в поведении обезьян. Одни животные сосредоточены на задаче и показывают стабильные результаты, другие животные менее внимательны и часто отвлекаются, что приводит к увеличению общего времени ответа.
Для объективной оценки индивидуальных особенностей поведения обезьян использовали метод измерения времени реакции на незнакомый предмет, отражающий выраженность исследовательского инстинкта [Coleman et al., 2005]. Животным последовательно предъявляли пять предметов и фиксировали время до первого касания. Обезьяны Н8 и Л8 не проявили особого интереса к новому: Л8 не взял ни одного предмета за три минуты, Н8 дотронулся лишь до одного предмета, предъявленного последним. Остальные животные исследовали по три–четыре предмета, но через разные промежутки времени. Так, животные Ф6 и Ю9 обследовали одинаковое число предметов (по три штуки), но Ф6 на это потребовалось в среднем более 30 с, а Ю9 справился с задачей за три секунды. Больше всех предметов обследовал Ф9: четыре штуки, но в среднем время до первого касания у него составило 42 с из-за того, что он долго не решался дотронуться до предмета № 4.
Для того чтобы учесть оба параметра: число обследованных предметов и время до касания предмета, был предложен способ перевода касаний в относительные единицы. Каждое касание оценивали как разность между максимальным временем ожидания (180 с) и фактическим временем в секундах, потребовавшимся обезьяне, чтобы дотронуться до предмета. Отсутствие ответной реакции принимали за ноль. Результаты реакции на каждый из пяти предметов суммировали и получили усредненный показатель исследовательской активности у каждой обезьяны, выраженный в относительных единицах (рис. 5 (а)).
Рис. 5.
Оценка исследовательской активности у обезьян. (а) – выраженность исследовательской активности у обезьян, рассчитанная на основе анализа реакции на незнакомые предметы. Особь Л8, как и остальные, участвовала в выполнении теста “реакция на незнакомый объект”, но не коснулась ни одного предмета, поэтому индекс исследовательской активности этой обезьяны равен 0; (б) – взаимосвязь уровня исследовательской активности и скорости обучения выполнению когнитивных тестов.
Fig. 5. Monkeys’ investigatory activity. (a) – the level of investigatory activity of the monkeys calculated from the reaction to novel objects test. Monkey Л8, together with the others, performed the novel object test but didn’t touch any items so its investigatory activity index was equal to 0; (б) – correlation between the level of investigatory activity and the cognitive task learning rate.
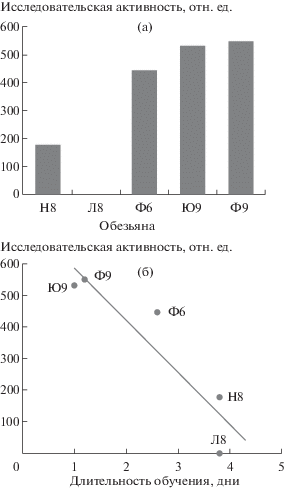
Как видно на рис. 5 (а), исследовательская активность наиболее выражена у обезьян Ю9 и Ф9. Чуть ниже этот показатель у обезьяны Ф6. Животные Н8 и Л8 оказались наименее заинтересованными в исследовании новых для них предметов. Подобное разделение животных мы наблюдали при анализе скорости обучения (рис. 2 (б)) – наиболее способными к обучению оказались обезьяны Ю9 и Ф9, дольше всех обучались обезьяны Л8 и Н8, животное Ф6 показало промежуточный результат.
Для того чтобы оценить взаимосвязь между скоростью обучения и выраженностью исследовательского инстинкта, мы рассчитали корреляцию между этими двумя показателями. Коэффициент корреляции Пирсона оказался статистически достоверным (p = 0.027, n = 5) и составил –0.92. Рис. 5 (б) иллюстрирует характер выявленной взаимосвязи: чем выше исследовательская активность, тем быстрее животное обучается выполнению когнитивных задач. Таким образом, анализ реакции обезьян на новые предметы можно использовать как экспресс-тест для выявления перспективных в плане обучения животных.
ОБСУЖДЕНИЕ РЕЗУЛЬТАТОВ
В результате проведенных исследований показана возможность обучения обезьян макак-резусов выполнению когнитивных тестов различной степени сложности, предъявляемых с помощью компьютера на сенсорном экране. Несмотря на существенные различия в скорости обучения, все животные после многократных тренировок успешно справлялись с поставленными задачами, включая такие, решение которых вызывает затруднения даже у человека. На рис. 6 приведено сравнение зависимостей количества правильных ответов от сложности теста, полученных на человеке и обезьяне с использованием одинаковых наборов матриц.
Рис. 6.
Сравнение результативности выполнения человеком и обезьяной одинаковых тестов. Результаты исследований на человеке взяты из статьи [Harauzov et al., 2016]. Жирные кривые на графиках – средние значения по всем испытуемым. Планки погрешности на правом графике – ошибка среднего.
Fig. 6. The comparison of the results obtained by humans and monkeys while performing the same task. The results of the experiments with human subjects are from the paper [Harauzov et al., 2016]. Thick lines are the mean values over all subjects. Error bars on the right plot are the error of mean.
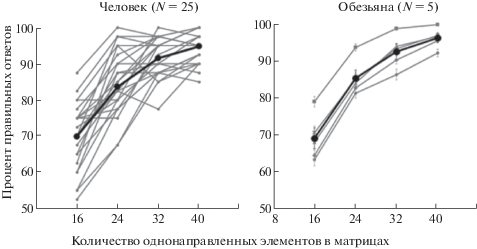
Как видно на рисунке, обезьяны не уступают человеку, решая самые сложные задачи даже лучше некоторых людей. Причем одна обезьяна решала все задачи выше среднего уровня правильных ответов, полученного на человеке. В среднем число правильных ответов у обезьян и людей было одинаково и составило 85%. Справедливости ради следует отметить, что у человека времени на раздумья было меньше: матрицы предъявляли на 1 с, и была инструкция дать ответ как можно скорее. У обезьян, напомним, ограничений во времени не было. В результате среднее время реакции на все типы матриц у человека составило 0.6 с, у обезьяны – 1.8 с. Этим, вероятно, объясняется отсутствие у большинства обезьян, в отличие от людей, зависимости времени ответа от сложности задачи.
Сходство кривых результативности человека и обезьяны позволяет предположить, что у обоих видов задействованы одинаковые нейронные механизмы при решении данного типа задач. Предположительно, при выполнении тестов с матрицами, как и многих других зрительных тестов, последовательно реализуются следующие нейронные механизмы. На начальном этапе происходит первичное описание физических характеристик стимулов, таких как яркость, контраст, пространственная частота и ориентация отдельных элементов. Затем отдельные фрагменты изображения должны быть объединены в единое целое. После этого происходит выделение доминирующей ориентации, предположительно, за счет нейронного механизма согласованной фильтрации [Красильников и др., 1999; Дудкин, 2007], который помогает выделить искомый признак путем сравнения информации, поступающей из внешнего мира, с шаблоном, или эталоном, хранящимся в памяти. Затем должны произойти сравнение двух стимулов и принятие решения, за которым следует развитие моторной реакции. Уточнить временную последовательность развития этих событий помогает регистрация вызванных потенциалов у человека [Хараузов и др., 2007; Сельченкова и др., 2011; Harauzov et al., 2016]. Возможность регистрации электрической активности глубинных структур мозга у обезьян в процессе выполнения тестов открывает широкие перспективы по изучению принципов действия, последовательности активации и локализации описанных нейронных механизмов. Полученные нами в поведенческих экспериментах схожие результаты выполнения человеком и обезьяной когнитивных задач позволят в дальнейшем экстраполировать на человека результаты электрофизиологических исследований с участием приматов, что, в свою очередь, открывает широкие перспективы использования последних в качестве животной модели изучения нейронных механизмов зрительного восприятия и их нарушения в результате различных патологий.
Анализ индивидуальных особенностей поведения обезьян показал, что выраженность исследовательской активности может значительно различаться от особи к особи. В литературе животных, которые с готовностью принимаются изучать новый объект, называют “исследователи” [Coleman et al., 2005; Fox, 1972; MacDonald, 1983] или “смелые” [Wilson et al., 1993]. Животных, которые относятся к новому стимулу с боязнью и не спешат вступать во взаимодействие с ним – “заторможенные” [Coleman et al., 2005; Boyce et al., 1998; Kagan et al., 1994; Williamson et al., 2003], “реактивные” [Suomi, 1988] или “застенчивые” [Wilson et al., 1993]. Обнаруженная в наших экспериментах высокая корреляция между уровнем исследовательской активности и скоростью обучения подтверждает результаты, сделанные в работе Coleman и соавторов [Coleman et al., 2005]: “исследователи” успешнее обучались выполнению простой когнитивной задачи (прикосновение к целевому объекту), чем “заторможенные” обезьяны. В указанной работе, однако, использовали всего один предмет и делили обезьян на три группы по времени до касания до него. В наших экспериментах число предметов было увеличено до пяти, и учитывали не только время до касания, но и число обследованных предметов. Кроме того, скорость обучения в наших исследованиях представляла интегрированный показатель, учитывающий способность к освоению нескольких однотипных задач, различающихся по степени сложности. Это увеличивает точность измерений и позволяет более тонко дифференцировать обезьян по уровню их исследовательской активности и скорости обучения.
Обезьяны с высоким уровнем исследовательской активности, помимо хорошей обучаемости, показывали более стабильное время ответа. Другими словами, эти обезьяны были больше мотивированы к работе и меньше отвлекались. Обезьяны с низким исследовательским индексом, наоборот, демонстрировали большое число так называемых уходов и могли нажать на экран через несколько десятков секунд. Такая особенность поведения, скорее всего, определяет скорость обучения. Чем выше мотивация к работе и устойчивость внимания, тем быстрее обезьяны осваивают новые задачи. Наши данные показывают, что все эти черты характера (поведенческие особенности) могут быть предсказаны с помощью теста “реакция на новый объект”.
Таким образом, измерение исследовательской активности по реакции на незнакомые предметы является простым и быстрым способом для отбора наиболее перспективных животных в плане обучения когнитивным задачам. Надежность способа повышается, если сравнивать обезьян, содержащихся в одинаковых условиях, поскольку, как показывают исследования [Основы зоопсихологии, 2001; Ерофеев и др., 2002], развитие исследовательского поведения, стрессоустойчивость и, следовательно, способность к обучению в довольно значительной степени определяются условиями, в которых растет детеныш, особенно возможностями общения с матерью, другими сородичами и особями иных видов. Учитывая, что обучение обезьян выполнению новых тестов зачастую требует больших временных затрат, в особенности, в эпоху компьютерных технологий, когда эксперименты становятся все сложнее, предварительный отбор особей при помощи конкретного метода поможет существенно сократить время экспериментаторов, затраченное на обучение обезьян новым задачам.
ВЫВОДЫ
1. В результате проведенных исследований показана возможность обучения обезьян Macaca mulatta выполнению пространственных когнитивных тестов с регулируемым уровнем сложности, предъявляемых на сенсорном мониторе компьютера.
2. В ходе выполнения когнитивных задач обезьяны продемонстрировали результаты, сходные с данными, полученными ранее на человеке. Это может позволить экстраполировать на человека результаты электрофизиологических исследований с участием приматов и использовать последних в качестве животной модели изучения нейронных механизмов зрительного восприятия и их нарушения в результате различных патологий.
3. Оценка исследовательской активности по реакции на незнакомые предметы (тест “реакция на новый объект”) может использоваться в качестве экспресс-методики для отбора наиболее перспективных животных с целью их последующего обучения когнитивным задачам.
Работа проведена на животных из ЦКП Биоколлекция ИФ РАН, поддержанной Программой ФАНО России по сохранению и развитию биоресурсных коллекций.
Работа выполнена при финансовой поддержке Программы фундаментальных научных исследований государственных академий на 2013–2020 годы (ГП-14, раздел 63).
Авторы благодарят И.В. Дворецкую, И.В. Капаеву, Д.А. Лучкина за помощь в проведении исследования.
Список литературы
Альтман Я.А. Слуховая система. Л.: Наука, 1990. 620 с.
Бонгард М.М. Проблема узнавания. М.: Наука, 1967. 320 с.
Дудкин К.Н. И.П. Павлов и нейрофизиология познавательных процессов. СПб: Ин-т физиологии им. И.П. Павлова РАН, 2007. 295 с.
Ерофеев А.И., Беляева Г.С., Тихонравов Д.Л. Особенности адаптации к экспериментальным условиям у приматов с различными исходными способами содержания. Ж. высш. нервн. деят. им. И.П. Павлова. 2002. 52 (6): 695–704.
Иванова Л.Е., Коржанова З.Н., Варовин И.А., Пронин С.В., Хараузов А.К., Шелепин Ю.Е. Изучение взаимодействия макак-резусов с тактильными мониторами при наблюдении низкочастотных тестовых изображений. Рос. физиол. журн. им. И.М. Сеченова. 2016. 102 (8): 931–939.
Коган А.Б. Основы физиологии высшей нервной деятельности. 2-е изд., перераб. и доп. М.: Высшая школа, 1988. 368 с.
Красильников Н.Н., Шелепин Ю.Е., Красильникова О.И. Фильтрация в зрительной системе человека в условиях порогового наблюдения. Оптический журнал. 1999. 66 (1): 5–14.
Основы зоопсихологии: Учебник для студентов высших учебных заведений, обучающихся по специальностям “Психология”, “Биология”, “Зоология” и “Физиология”. 3-е изд. М.: Учебно-методический коллектор “Психология”, 2001. 464 с.
Сельченкова Т.В., Хараузов А.К., Шелепин Ю.Е. Электрофизиологические и психофизические исследования влияния длительности предъявления текстур на пороги распознавания. Рос. физиол. журн. им. И.М. Сеченова. 2011. 97 (3): 316–329.
Стреляу Я. Роль темперамента в психическом развитии. М.: Прогресс, 1982. 232 с.
Хараузов А.К., Шелепин Ю.Е., Пронин С.В., Сельченкова Т.В., Носков Я.А. Электрофизиологическое исследование механизмов распознавания текстур. Рос. физиол. журн. им. И.М. Сеченова. 2007. 93 (1): 3–13.
Шелепин Ю.Е., Фокин В.А., Хараузов А.К., Пронин С.В., Чихман В.Н. Локализация центра принятия решений при восприятии формы зрительных стимулов. Докла. акад. наук. 2009. 429 (6): 1–3.
Boyce W.T., O’Neill-Wagner P., Price C.S., Haines M., Suomi S.J. Crowding stress and violent injuries among behaviorally inhibited rhesus macaques. Health Psychol. 1998. 17: 285–289.
Coleman K., Tully L.A., McMillan J.L. Temperament Correlates with Training Success in Adult Rhesus Macaques. Am. J. of Primatol. 2005. 65: 63–71.
Crawford M.P. A behavior rating scale for young chimpanzees. Journ. of Compar. Psychol. 1938. 26: 79–91.
De Valois R.L., Morgan H., Snodderly D.M. Psychophysical studies of monkey vision-III. Spatial luminance contrast sensitivity tests of macaque and human observers. Vision Res. 1974a. 14: 75–81.
De Valois R.L., Morgan H.C., Polson M.C., Mead W.R., Hull E.M. Psychophysical studies of monkey vision–I. Macaque luminosity and color vision tests. Vision Res. 1974b. 14: 53–67.
Fox M.W. Socio-ecological implications of individual differences in wolf litters: a developmental and evolutionary perspective. Behavior. 1972. 41: 298–313.
Freeman H.D., Gosling S.D. Personality in nonhuman primates: a review and evaluation of past research. Am. J. of Primatol. 2010. 72 (8): 653–671.
Griffin A.S., Guillette L.M., Healy S.D. Cognition and personality: an analysis of an emerging field. Trends in Ecology and Evolution. 2015. 30 (4): 207–214.
Harauzov A.K., Shelepin Y.E., Noskov Y.A., Vasilev P.P., Foreman N.P. The time course of pattern discrimination in the human brain. Vision Res. 2016. 125: 55–63.
Heath-Lange S., Ha J.C., Sackett G.P. Behavioral measurement of temperament in male nursery-raised infant macaques and baboons. Am. J. of Primatol. 1999. 47 (1): 43–50.
Hebb D.O. Emotion in man and animal: An analysis of the intuitive process of recognition. Psychol. Rev. 1946. 53: 88–106.
Herrmann E., Call J., Hernández-Lloreda M.V., Hare B., Tomasello M. Humans have evolved specialized skills of social cognition: the cultural intelligence hypothesis. Science. 2007. 317: 1360–1366.
Kagan J., Reznick J., Snidman N. Temperamental variation in response to the unfamiliar. Perinatal development: a psychobiological perspective. Ed. Krasnegor N., Blass E., Hofer M., Smotherman W. Orlando, FL: Academic Press, 1987. 421–440 pp.
MacDonald K. Stability of individual differences in behavior in a litter of wolf cubs (Canis lupis). J. Comp. Psychol. 1983. 97: 99–106.
NIST/SEMATECH e-Handbook of Statistical Methods http://www.itl.nist.gov/div898/handbook/
Passingham R. How good is the macaque monkey model of the human brain? Curr. Opin. Neurobiol. 2009. 19 (1): 6–11.
Rajalingham R., Schmidt K., DiCarlo J.J. Comparison of Object Recognition Behavior in Human and Monkey. J. of Neurosc. 2015. 35 (35): 12127–12136.
Réale D., Reader S.M., Sol D., McDougall P.T., & Dingemanse N.J. Integrating animal temperament within ecology and evolution. Biol. Rev. Camb. Philos. Soc. 2007. 82 (2): 291–318.
Schmitt V., Kröger I., Zinner D., Call J., Fischer J. Monkeys perform as well as apes and humans in a size discrimination task. Anim. Cogn. 2013. 16 (5): 829–838.
Suomi S.J. Genetic and maternal contributions to individual differences in rhesus monkey biobehavioral development. Psychobiological aspects of behavioral development. Ed. Krasnagor N. New York: Academic Press, 1988. 397–419 pp.
Va’zquez P., Cano M., Acun˜a C. Discrimination of line orientation in humans and monkeys. J. Neurophysiol. 2000. 83: 2639–2648.
Williamson D.E., Coleman K., Bacanu S.A., Devlin B.J., Rogers J., Ryan N.D., Cameron J.L. Heritability of fearful-anxious endophenotypes in infant rhesus macaques: a preliminary report. Biol. Psychiatry. 2003. 53: 284–291.
Wilson D.S., Coleman K., Clark A.B., Biederman L. The shy–bold continuum in pumpkinseed sunfish (Lepomis gibbosus): an ecological study of a psychological trait. J. Comp. Psychol. 1993. 107: 250–260.
Yerkes R.M. The life history and personality of the chimpanzee. The American Naturalist. 1939. 73: 97–112.
Дополнительные материалы отсутствуют.
Инструменты
Журнал высшей нервной деятельности им. И.П. Павлова