

Исследование Земли из Космоса, 2020, № 2, стр. 35-55
Моделирование распределения потенциальных местообитаний переднеазиатского леопарда (Panthera pardus ciscaucasica) на Северном Кавказе методом MaxEnt на основе данных спутникового мониторинга выпущенных в природу животных
В. В. Рожнов a, *, Р. Х. Пшегусов b, Х. А. Эрнандес-Бланко a, М. Д. Чистополова a, А. Б. Пхитиков b, С. А. Трепет b, c, Н. А. Дронова d, С. В. Найденко a, А. А. Ячменникова a
a Институт проблем экологии и эволюции им. А.Н. Северцова Российской академии наук
Москва, Россия
b Институт экологии горных территорий им. А.К. Темботова Российской академии наук
Нальчик, Россия
c Кавказский государственный природный биосферный заповедник им. Х.Г. Шапошникова
Сочи, Россия
d Всемирный фонд дикой природы, ВВФ России
Москва, Россия
* E-mail: rozhnov.v@gmail.com
Поступила в редакцию 03.12.2019
Аннотация
В рамках программы по восстановлению леопарда на Кавказе в 2016 и 2018 гг. были выпущены в природу первые леопарды, меченые GPS-ошейниками со спутниковыми передатчиками. Данные, полученные с ошейников, были использованы для создания и верификации математической модели потенциальных местообитаний этого вида и оценке возможного использования леопардами. Моделирование выполнено методом максимальной энтропии с использованием программы MaxEnt на основе данных по перемещению шести леопардов (четыре самца и две самки). Разнообразие основных свойств местообитаний отражено в биоклиматических показателях WorldClim, данных радиолокационной съемки поверхности и индексах, рассчитанных на основе мозаики спутниковых снимков Landsat8. Для описания и верификации биотопов использовали полевые данные, собранные на территории Республик Северная Осетия-Алания, Южная Осетия и Кабардино-Балкария. Анализ результатов моделирования показал высокую точность определения предсказанных точек. Из всего комплекса факторов (115) наибольший вклад в формирование общей модели потенциальных местообитаний леопардов вносят 10, среди которых крутизна склона, высота над уровнем моря, ряд климатических характеристик, биоклиматические индексы и вегетационный индекс. По результатам моделирования, наиболее предпочтительные для леопарда местообитания на Кавказе представляют собой не очень крутые (до 30°) склоны с травянистой растительностью на высотах порядка 800–1500 м н.у.м., не зависящие от экспозиции, но обладающие средними значениями снеговых индексов.
ВВЕДЕНИЕ
Сохранение и восстановление флаговых редких видов животных представляет собой комплексную фундаментально-прикладную задачу, позволяющую, в том числе, обеспечивать сохранение экосистем в целом. Реинтродукция животных является одним из важнейших методов сохранения и восстановления численности видов, находящихся под угрозой исчезновения (Sarrazin, Barbault, 1996; Hayward, Somers, 2009). Общепризнанной практикой является создание центров разведения животных и их подготовки к выпуску в природу c последующей реинтродукцией в пределах их естественного ареала. Основные требования к местам выпуска следующие: они должны быть расположены в пределах исторически сформировавшегося ареала, иметь хорошую кормовую базу, находиться вдали от поселений человека (Hayward, Somers, 2009). Исторически обитавший на Кавказе переднеазиатский леопард (Panthera pardus ciscaucasica) является подвидом с самыми крупными представителями в мире и в настоящее время занесен в Приложение I Конвенции о международной торговле видами дикой фауны и флоры, находящимися под угрозой исчезновения (CITES). В 2007 году Институтом проблем экологии и эволюции им. А.Н. Северцова Российской академии наук (ИПЭЭ РАН) была разработана Программа восстановления популяции переднеазиатского леопарда на Кавказе путем разведения его в неволе и последующей реинтродукции животных, соответствующих характерным для диких особей нормам поведения и физиологическим статусом, (Программа…, 2008). В 2016 г. на Западном Кавказе и 2018 г. на Западном и Центральном Кавказе выпустили первых леопардов, подготовленных к жизни в дикой природе в специализированном Центре восстановления леопардов на Кавказе, на базе Сочинского национального парка (Семенов, 2016). Перед выпуском животных снабдили ошейниками с GPS-приемником и спутниковым передатчиком (Рожнов и др., 2018; Rozhnov et al., 2019). Координаты локаций, полученные с ошейников выпущенных леопардов, позволяют выявить процесс освоения пространства и использование территории после выпуска в природу.
В настоящее время все более актуальными при работе с редкими видами становятся неинвазивные методики разных категорий (Рожнов и др., 2018; Rozhnov et al., 2019). К таким методам относят и математическое моделирование вариантов развития судьбы популяций (Kanagaraj et al., 2013; Carter et al., 2015; Гольцман и др., 201…, Родникова и др., 2018), вариантов распределения и местообитаний на основе характеристик среды (Safford, 2004; Hebblewhite et al., 2014; Добрынин и др., 2017; Жу, 2019; Rozhnov et al., 2019). Существуют многочисленные алгоритмы моделирования для прогнозов пространственного распределения видов и предположительной оценки площадей их местообитаний (Stockwell, Peters, 1999; Guisan, Zimmermann, 2000; Austin, 2002; Guisan et al., 2007; Guo, Liu 2010; Hijmans et al., 2005; Stigall, 2012). Математический алгоритм максимальной энтропии (Phillips et al., 2006; Phillips, Dudik, 2008; Elith et al., 2010), встроенный в программное обеспечение MaxEnt (виды максимальной энтропии при моделировании распределения), в настоящее время распространен наиболее широко. Работа алгоритма основана только на данных достоверного присутствия вида. А аппроксимация удовлетворяет всем ограничениям, известным исследователю, при этом результирующее распределение имеет максимальную энтропию (Phillips et al., 2006). Результат вычислений алгоритма выглядит как карта с прогнозами вероятности присутствия вида в каждой ячейке/пиксельной сетке. Одной из частных проблем изучения экологии животных в условиях горных территорий является труднодоступность большинства участков для прямой оценки исследователями. Решение данной проблемы, в том числе, относится к полю неинвазивных методов – дистанционного зондирования и интерполяции результатов, полученных для ограниченной территории, на площадь всего региона. Результаты пространственного моделирования полезны: при планировании работ, как по освоению (Safford, 2004), так и по сохранению (Rondinini et al., 2005) окружающей среды; при ретроспективном анализе эволюционных процессов в пределах территорий расселения популяций; при управлении инвазивными видами – регулировании их плотности; для иных целей, включая определение пригодности среды обитания вида (Jiang et al., 2016), прогнозирование тенденций распространения видов на региональном уровне (Giovannelli et al., 2010), а также для выявления потенциальных зон высокого риска, зон вероятного скорого конфликта между дикой природой и человеком (Naqibzadeh et al., 2019).
Целью данной работы было охарактеризовать места, которые могут быть выбраны с высокой долей вероятности леопардами для обитания или перемещения на Западном и Центральном Кавказе, определить характеристики таких мест на основе вычислений моделирования, базирующегося на реальных данных, полученных от ошейников выпущенных леопардов и ДДЗЗ, а также на результатах полевой верификации биотопов в одном из модельных регионов (территория республик Северная Осетия-Алания, Южная Осетия и Кабардино-Балкария).
В рамках поставленной цели были определены две основные задачи. Первая – оценить вероятность использования местообитаний с различными характеристиками на основе совокупности данных, полученных от ошейников выпущенных животных на наиболее обширной площади Западного и Центрального Кавказа. Вторая – проверить предсказательную способность модели в три этапа (до выпуска животных; через полгода после выпуска, через год после выпуска), учитывая плотность населения и полевую верификацию местообитаний. В качестве объекта для проверки предсказательной способности модели выбрали одну самку леопарда по кличке Волна и территории субъектов Кавказского региона – Кабардино-Балкарскую Республику и Республику Северная Осетия-Алания.
МАТЕРИАЛ И МЕТОДЫ
Район исследований
В ходе работы моделирование потенциально пригодных местообитаний переднеазиатского леопарда и интерполяцию полученных результатов осуществляли для территории, охватывающей Кавказскую горную страну в следующих физико-географических границах: на востоке – Каспийское море, на севере – от Каспийского моря до Кумо-Манычской впадины и далее по границе Краснодарского края до Черного моря, на западе – по побережью Черного моря, на юге – по Кура-Аракскинской низменности и южным границам Азербайджана, Армении и Грузии (рис. 1).
Основа моделирования – GPS-локации
В качестве базы в ходе построения модели использовали координаты точек, полученных в результате спутникового GPS-мониторинга шести выпущенных переднеазиатских леопардов: 2 самок (Виктория, Волна), 4 самцов (Ахун, Килли, Артек и Эльбрус). Самка Виктория и два самца (Килли и Ахун) были выпущены в 2016 году на Западном Кавказе в Кавказском заповеднике (Краснодарский край, Карачаево-Черкесия, Адыгея), там же в 2018 году был выпущен самец Артек. Самка Волна и самец Эльбрус были выпущены в 2018 году на Центральном Кавказе на территории Национального парка “Алания” (Республика Северная Осетия). Леопарды были снабжены ошейниками с GPS-приемниками и спутниковыми передатчиками Lotek GPS Iridiumtrack M (LOTEK Engineering Ltd., Ньюмаркет, ON, Канада). Ошейники леопардов, выпущенных в 2016 г., были запрограммированы на определение местоположения каждые два часа (12 локаций в сутки) и передачу накопленных данных один раз в сутки. Ошейники леопардов, выпущенных в 2018 г., определяли местоположение каждый час (24 локации в сутки) и передачу два раза в сутки. Количество полученных локаций для каждого леопарда со времени выпуска до даты прекращения работы передатчика приведено в табл. 1. Всего в моделировании использовано 14620 точек. При оценке возможного пространственного распределения леопарда по всей территории Кавказа были проанализированы потенциально пригодные местообитания для вида в целом. Также рассмотрены перспективы использования пространства животными раздельно по половому признаку и для каждой особи индивидуально.
Таблица 1.
Общие характеристики проведенного моделирования потенциальных местообитаний: основа для построения модели в виде количества локаций, точность полученной модели, вычисленная предсказанная площадь
| Особи леопарда, по чьим данным рассчитана модель | Численные показатели | ||
|---|---|---|---|
| N (количество точек локаций, включенных в анализ) | AUC ± SDauc | площадь, км2 | |
| Все животные (All) | 14 620 | 0.935 ± 0.003 | 4508.13 |
| Все самки (♀♀Female) | 6416 | 0.977 ± 0.002 | 705.64 |
| Все самцы (♂♂Male) | 8204 | 0.944 ± 0.002 | 2365.04 |
| Ахун (♂Ahun) | 214 | 0.988 ± 0.001 | 82.50 |
| Артек (♂Artek) | 2989 | 0.969 ± 0.001 | 983.96 |
| Эльбрус (♂Elbrus) | 1709 | 0.981 ± 0.002 | 808.15 |
| Килли (♂Killi) | 3292 | 0.970 ± 0.002 | 1190.76 |
| Виктория (♀Viktoria) | 3048 | 0.986 ± 0.001 | 610.49 |
| Волна (♀Volna) | 3368 | 0.9981 ± 0.000 | 738.09 |
Параметры моделирования
Для построения интерполяционной модели были использованы характеристики среды – 115 переменных, объединенных в три блока.
1. Биоклиматические показатели WorldClim, включающие среднемесячные данные по средней, минимальной и максимальной температуре, осадкам, солнечной радиации, давлению водяного пара, скорости ветра – 7 переменных среды по 12 месяцам (всего 84) и производные от них биоклиматические характеристики – 19 биоклиматических параметров (Hijmans et al., 2005), обозначенные в работе следующим образом: tmin (1–12) – минимальные значения среднемесячной температуры, °C; tmax (1–12) – максимальные значения среднемесячной температуры, °C; tavg (1–12) – среднемесячная температура, °C; prec (1–12) – количество осадков, мм; srad (1–12) – солнечная радиация, кДж/м2 в день; wind (1–12) – скорость ветра, м/с; vapr (1–12) – давление водяного пара, кПа; bio1 – среднегодовая температура, °C; bio2 – среднесуточная амплитуда температур, °C; bio3 – изотермальность, отношение среднесуточной и среднегодовой температур, %; bio4 – температурная сезонность, стандартное отклонение температуры, %; bio5 – максимальная температура наиболее теплого месяца, °C; bio6 – минимальная температура наиболее холодного месяца, °C; bio7 – годовая амплитуда температур, °C; bio8 – средняя температура наиболее влажного квартала, °C; bio9 – средняя температура наиболее сухого квартала, °C; bio10 – средняя температура наиболее теплого квартала, °C; bio11 – средняя температура наиболее холодного квартала, °C; bio12 – годовые осадки, мм; bio13 – количество осадков в наиболее влажный месяц, мм; bio14 – количество осадков в наиболее сухой месяц, мм; bio15 – сезонность осадков, коэффициент вариации; bio16 – количество осадков в наиболее влажный квартал, мм; bio17 – количество осадков в наиболее сухой квартал, мм; bio18 – количество осадков в наиболее теплый квартал, мм; bio19 – количество осадков в наиболее холодный квартал, мм. Также в работе использованы переменные, характеризующие снежный покров – snowcover (кг/м2), глубину снега – snowdepth (м), основанные на глобальной базе климатических данных – Worldclim2 и FLDAS Noah Land Surface Model (McNally et al., 2017) – всего 2 показателя снежного покрова;
2. Данные радиолокационной съемки рельефа Global Multi-resolution Terrain Elevation Data 2010 (GMTED2010) с разрешением 250 м. На их основе рассчитывали высоту над уровнем моря (alt) и морфометрические характеристики рельефа: уклон – slope, экспозиция – aspect, различные виды кривизны (общая – curv, плановая – plcurv, продольная – longcurv и поперечная – cscurv), показатели освещенности склонов с востока (sh_r_e) и юга (sh_r_s) и индекс Лапласиана (lap): 10 морфометрических характеристик уклон, экспозиция, различные виды кривизны (общая, плановая, продольная и поперечная), показатели освещенности склонов с востока и юга и индекс Лапласиана: 10 морфометрических характеристик рельефа (Zevenbergen, Thorne, 1987; Moore et al., 1991; Buckley, 2010), которые в горах могут непосредственно влиять на размещение видов;
3. Вегетационные индексы NDVI летнего (ndvi_s) и зимнего (ndvi_w) периода, рассчитанные на основе измерения отраженной солнечной радиации мозаики территории исследования со спутника Landsat 8OLI/TRS – 2 вегетационных индекса.
Вопрос использования коррелированных факторов в моделях, проблема коллинеарности и увеличения неопределенности параметров, рассматриваемые в обширной литературе, посвященной моделированию методом MaxЕnt, остается дискуссионным (Dormann et al., 2008; Araujo, Peterson, 2012; De Marco, Nobrega, 2018). В нашей работе приводится результат моделирования, с использованием всей доступной информации: 115 переменных, приведенных выше. Все использованные в работе переменные приведены к разрешению 250 м/пикс. программным методом би-кубической интерполяции сплайнами (Studley, Weber, 2011).
Метод моделирования
Определение потенциально возможных местообитаний проводили методом максимизации энтропии (Maxent software for species habitat modeling). Данный метод является сравнительно новым, но он стал популярным в последние годы подходом в изучении распространения животных (Baldwin, 2009; Elith et al., 2006, 2010; Огурцов, 2019). Он хорошо зарекомендовал себя при исследовании и прогнозировании пространственного распределения различных компонентов горных экосистем Кавказа: лишайников и печеночников (Пшегусов, Ханов, 2015; Жашуев, Пшегусов, 2016), травяных сообществ (Цепкова и др., 2015), хищных птиц и крупных млекопитающих (Пшегусов, 2014; Bleyhl et al., 2015). Методы MaxЕnt в соответствие с их логико-математической основой и использованием информации только о наличии вида выделяют наиболее оптимальные местообитания. При этом возможна неоднократная оценка и верификация модели. Для получения наиболее объективной итоговой модели в ходе данного исследования провели 15 репликаций (повторных вычислений) по 500 итераций в каждом пикселе всей анализируемой области, 25% дискретных точек в каждом анализе случайным образом были определены предварительно как проверочные.
Результаты выведены с оценкой вероятности присутствия вида в диапазоне от 0 до 1. Области с вероятностью встречаемости вида 0,8 и выше рассматриваются нами как оптимальные (такие, где особи вида могут быть обнаружены с вероятностью 80% и выше). Области с вероятностью от 0,5 до 0,8 – охарактеризованы как потенциально пригодные, т.е. это области, которые вид может занять при определенных условиях или использовать для перемещения, расселения, во время миграционных переходов.
Верификация модели
Одной из важных задач в ходе прогнозирования потенциально пригодных территорий является верификация модели. Стандартно данная процедура производится сравнением значений потенциальной пригодности местообитаний, смоделированной в MaxЕnt, с фактическими данными, собираемыми в полевых исследованиях, в том числе и по результатам моделирования (West et al., 2016). В данной работе верификацию осуществляли по локациям, полученным от ошейников выпущенных леопардов. Был апробирован подход, с помощью которого на примере перемещений самки Волна мы провели анализ значений вероятности встречаемости в точках локаций с применением инструмента геообработки Extract Multi Values To Points в расширении ArcGIS Spatial Analyst. Всего верификация проходила в три этапа. На первом, предварительном этапе (до выпуска в природу), был проведен расчет модели и оценка территории, которую может выбрать леопард после выпуска. Данное моделирование основывали на информации о двух локациях – реальных наблюдениях встреч диких леопардов на территории Северной Осетии – регистрации на территориях Зарамагской (2015 и 2017 гг.) и Гизельдонской (2012 г.) ГЭС. В силу специфики объекта исследований и высокой степени случайности предсказанных точек, изначальная модель не характеризуется высокой достоверностью (AUC = 0.892 ± 0.003). Однако мы сочли целесообразным произвести ее расчет, поскольку работа по реинтродукции данного вида крупной кошки не имеет аналогов в мире, а дикие особи встречаются настолько редко в природе России, что каждая встреча может считаться уникальной. В связи с этим, даже вычисление модели такого характера может дать информацию для планирования работ по выпуску животных. На втором этапе была построена модель, основанная уже непосредственно на локациях ошейника, полученных за первые полгода использования пространства самкой после выпуска (Volna 2018; AUC = 0.932 ± 0.001). На третьем этапе построили модель по полному набору данных за весь период работы ошейника (Volna 2019; AUC = 0.998 ± 0.000). Также использовали результаты расчета полной модели по всем леопардам (см. выше) и модели, посчитанной отдельно для самок. В ходе анализа полный набор точек локаций самки Волна был наложен на каждую другую модель и в результате извлечения значения ячеек из одного или нескольких растров в класс точечных объектов получены значения вероятности по каждой точке. Результаты верификации приведены в соответствующем пункте раздела “Результаты и обсуждение”.
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Общая характеристика моделирования
Для исследуемого вида моделирование показало высокую точность (неслучайность) определения предсказанных точек. Показатель, представляющий площадь под операционной кривой (Area Under the Curve), используемый для оценки полученной модели, характеризуется высокими значениями (среднее значение AUC для 15 повторных моделей для вида в целом составляет 0.935, а стандартное отклонение – 0.003), значения для других расчетов приведены в табл. 1.
На основании результатов моделирования, с учетом различной площади территории, освоенной каждой особью (информация от передатчиков, установленных на ошейниках), определена площадь оптимальных для леопардов местообитаний (табл. 1). Так, согласно модели по совокупным данным со всех ошейников, общая площадь территории, где вероятность обитания леопардов наиболее высока, составляет порядка 4500 км2 (450 тыс. га) – рис. 2. Поскольку площадь, используемая одним животным этого вида, может варьировать от 15 до 45 тыс. га (Чистополова и др., 2018; Fahradinia et al., 2018) в зависимости от плотности популяции вида, от плотности и системы распределения жертв в пространстве, а также внешних условий, проанализированная территория может быть пригодна для обитания 5–7 особей леопарда (например, для двух размножающихся самок, одного самца и 3–4 разновозрастных неполовозрелых особей). Учитывая, что индивидуальные участки обитания леопардов (Swanepoel, Van Hoven, 2008; Чистополова и др., 2018), в том числе и переднеазиатских (Farhadinia et al., 2018), могут перекрываться, данная территория может вместить не менее 10 (до 30) особей, т.е. 5 (до 15) пар леопардов.
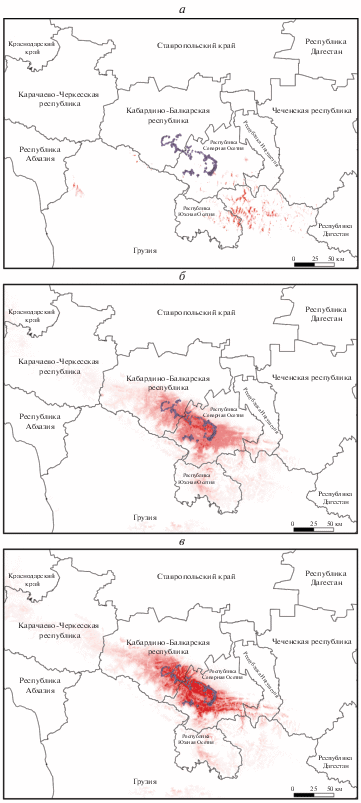
Рис. 2.
Модель потенциально пригодных местообитаний леопарда на Западном и Центральном Кавказе по данным спутниковых ошейников: а – все особи; б – самки; в – самцы. Градация шкалы: от 0 (белый, непригодные местообитания) до 1 (красный, оптимальные местообитания).
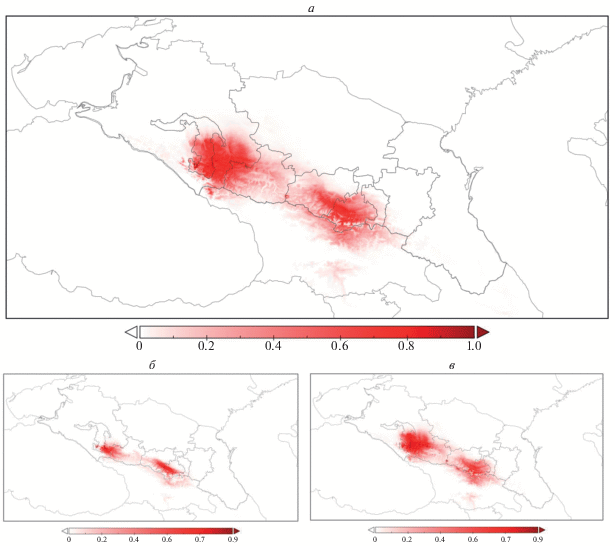
Из полученных результатов (рис. 2, 3) можно также предположить возможные направления расширения площади, используемой леопардами, в том числе пути переходов животных из Западно-кавказского региона в местообитания на Центральном Кавказе. Исходя из модели, определяются два “коридора”. Один из них, более высокогорный, предполагает, в том числе, некоторые участки южного макросклона Большого Кавказского хребта и охватывает также верхнюю границу лесного пояса с прилегающими выше субальпийскими и альпийскими ландшафтами. Другой расположен в зоне предгорий, ближе к границе лесных ценозов и предгорных открытых участков, том числе антропогенно трансформированных ландшафтов. Эти направления характерны для перехода с Западного Кавказа на Центральный (территорию Кабардино-Балкарии), через Карачаево-Черкесию. С Центрального Кавказа (территории Осетии) на Западный Кавказ переход один, он наиболее обширный и локализуется преимущественно в поясе широколиственных лесов. Поскольку в полосе предгорий плотность населения и расположения населенных пунктов наиболее высокая, то при возможном перемещении хищников наиболее вероятным представляется “коридор” вдоль верхней границы широколиственных лесов.
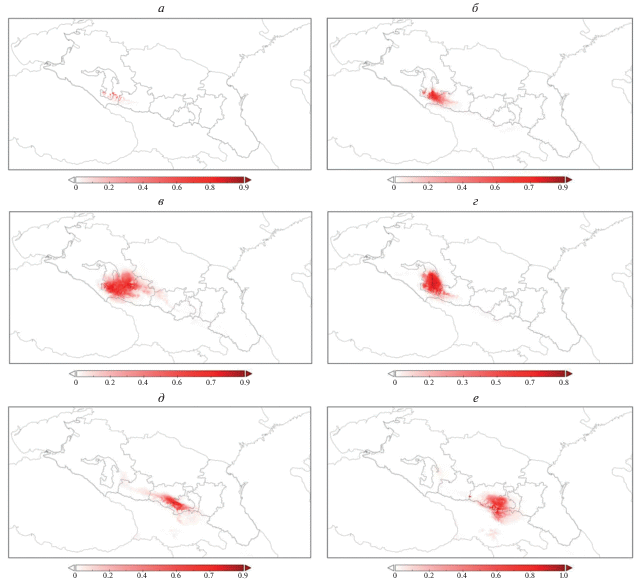
Рис. 3.
Модель потенциально пригодных местообитаний леопарда на Западном и Центральном Кавказе по данным спутниковых ошейников: особи, выпущенные на Западном Кавказе: а – самец Ахун, 2016 г. вып.; б – самка Виктория, 2016 г. вып.; в – самец Килли, 2016 г. вып.; г – самец Артек, 2018 г. вып. Особи, выпущенные на Центральном Кавказе: д – самка Волна, 2018 г.вып., е – самец Эльбрус, 2018 г. вып.
Анализ пространственного распределения леопарда по результатам моделирования
Из рис. 2 и 3 видно, что полученная модель затрагивает и южные склоны Большого Кавказа, а также возможные переходы с Большого Кавказа на Малый, соответствующие району Лихского (Сурамского) хребта. Этот естественный природный мост использовался леопардами для переходов в прошлом (до уничтожения человеком), о чем свидетельствуют многие исторические источники (Верещагин, 1959; Гептнер, Слудский, 1972). Соответствие прогнозных вариантов расширения ареала в Закавказье с указанными в литературе историческими местами регистрации этого вида также свидетельствует, на наш взгляд, о высокой точности модели.
Полученные данные (табл. 1, рис. 2) говорят о том, что самцы менее требовательны к условиям пространства, чем самки, и для них в большей степени характерно исследовательское поведение, в результате чего они осваивают бóльшие по площади территории, обладающие бóльшим спектром и разнообразием условий, чем самки. С этим предположением соотносятся результаты, приведенные в табл. 2 и 3а, б: группа факторов (характеристик пространства), наиболее значимых для модели пригодных для самцов территорий, значительно уже, чем группа факторов, определяющих выбор географического участка самкой. При этом в данной модели мы не учитывали информацию по плотности жертв, а также степень антропогенной нагрузки на регион, которая неравномерна. Также это соотносится с результатами оценки выбора пути животными в соответствии с рельефными направляющими (Рожнов и др., 2019). Крупные кошачьи зачастую предпочитают перемещаться вдоль склонов, ориентируясь, на естественные направляющие рельефа (Koshkarev, 1984). По нашим предварительным данным самки, в том числе и переднеазиатского леопарда, предпочитают наименее энергозатратную стратегию перемещения вдоль склонов, тогда как перемещение самцов характеризуется большим диапазоном высотных и рельефных предпочтений и перепадом высот (Рожнов и др., 2019).
Таблица 2.
Некоторые характеристики местообитаний с вероятностью встречаемости леопардов от 0.8 и выше
| Модель | Значение | Характеристики местообитаний | |||
|---|---|---|---|---|---|
| высота | экспозиция склона | крутизна склона | вегетационный индекс | ||
| Все леопарды | Среднее | 1374.34 | 169.61 | 8.51 | 0.75 |
| Минимум | 571.00 | 2.68 | 0.87 | 0.13 | |
| Максимум | 2814.00 | 355.05 | 27.01 | 0.83 | |
| Стандартное отклонение | 573.61 | 97.27 | 4.84 | 0.11 | |
| Коэффициет вариации | 41.74 | 57.35 | 56.89 | 14.96 | |
| Самцы | Среднее | 1189.54 | 197.58 | 7.21 | 0.76 |
| Минимум | 337.00 | 4.68 | 0.66 | 0.14 | |
| Максимум | 4810.00 | 353.05 | 27.01 | 0.84 | |
| Стандартное отклонение | 607.39 | 92.79 | 4.74 | 0.14 | |
| Коэффициет вариации | 51.06 | 46.96 | 65.61 | 17.41 | |
| Самки | Среднее | 1582.34 | 181.49 | 0.74 | 10.58 |
| Минимум | 623.00 | 5.71 | 0.36 | 1.98 | |
| Максимум | 3022.00 | 350.69 | 0.83 | 30.45 | |
| Стандартное отклонение | 456.61 | 99.82 | 0.08 | 5.43 | |
| Коэффициет вариации | 28.85 | 55.01 | 11.11 | 51.30 | |
Таблица 3
а. Вклад факторов среды в прогностическую модель встречаемости вида в целом и отдельно для самок и самцов
| Все леопарды (all) | Самки (female) | Самцы (male) | ||||||
|---|---|---|---|---|---|---|---|---|
| переменная (variable) |
вклад переменной (%) percent contribution |
значимость пермутации (permutation importance) | переменная (variable) |
вклад переменной (%) percent contribution |
значимость пермутации (permutation importance) | переменная (variable) |
вклад переменной (%) percent contribution |
значимость пермутации (permutation importance) |
| prec8 | 36.4 | 0.1 | prec1 | 25.9 | 13.8 | prec8 | 50.2 | 0 |
| alt | 13.1 | 3 | alt | 25.4 | 2 | wind3 | 11.5 | 16.7 |
| bio18 | 10.5 | 1.2 | bio18 | 13.9 | 0.3 | alt | 8.7 | 4.1 |
| wind3 | 9.8 | 13.7 | srad9 | 4.7 | 11.7 | prec1 | 7 | 0 |
| prec1 | 9.7 | 0.5 | wind3 | 4.2 | 0.9 | bio9 | 3.2 | 2.2 |
| srad9 | 3.6 | 11 | srad3 | 3.7 | 25.9 | ndvi_s | 2.6 | 0.5 |
| bio9 | 3.5 | 3.8 | bio17 | 3.7 | 0.3 | prec3 | 2.5 | 2.4 |
| prec10 | 3.2 | 4.4 | srad12 | 2 | 2.7 | bio18 | 2.4 | 1.2 |
| ndvi_s | 1.5 | 1.2 | wind7 | 2 | 4 | srad9 | 2.2 | 8.2 |
| prec5 | 1.1 | 30 | ndvi_w | 1.4 | 0 | prec10 | 1.5 | 2.1 |
| ndvi_s | 1.4 | 1.7 | ||||||
| bio15 | 1.2 | 5.2 | ||||||
| slope | 1.1 | 2.7 | ||||||
| prec10 | 1.1 | 3.8 | ||||||
Если рассматривать территории, наиболее предпочитаемые леопардом с точки зрения географии места выпуска, для животных, выпущенных в Кавказском заповеднике, набор факторов, определяющих местообитания несколько иной, чем у животных, выпущенных в Северной Осетии. Это, вероятно, может быть связано как с бóльшей вариабельностью и сравнительной мягкостью климатических условий в северо-западной части Кавказа относительно центральной его части, так и с меньшей заселенностью региона человеком. В каждой новой ситуации на выбор зверем местоположения влияет неодинаковый набор факторов на каждой территории. Так, в западной части Кавказа территории с благоприятным для леопарда и копытных рельефом более обширные, высоты не такие большие, как на Центральном Кавказе, лесопокрытая площадь больше, площадь субальпики также обширнее, климат мягче (Гвоздецкий,1963; Герасимов и др., 1980). Все это в сочетании с обширными площадями ООПТ (Особо охраняемые…, 2009) обуславливает, в том числе, и высокое разнообразие потенциальной кормовой базы леопарда на Западном Кавказе. На Центральном Кавказе условия более жесткие: диапазон критических значений внешних факторов (климат и рельеф) больше, чем на Западном Кавказе. Соответственно, ограниченное число факторов (как количественных, так и качественных), определяющих выбор леопардами территории на участках Центрального Кавказа, отличается от такового для Западного Кавказа. Вдобавок к этому площадь ненарушенных территорий на Центральном Кавказе меньше, а их мозаичность напротив выше (Krever et al., 2001).
Таблица 4
Таблица 3б. Вклад факторов среды в прогностическую модель встречаемости по особям
| Ахун (♂Akhun) | Артек (♂Artek) | Эльбрус (♂Elbrus) | Килли (♂Killi) | Виктория (♀Victoria) | Волна (♀Volna) | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| переменная | вклад переменной (%) | Значимость пермутации | переменная | вклад переменной (%) | Значимость пермутации | переменная | вклад переменной (%) | Значимость пермутации | переменная | вклад переменной (%) | Значимость пермутации | переменная | вклад переменной (%) | Значимость пермутации | переменная | вклад переменной (%) | Значимость пермутации |
| prec4 | 15.2 | 0 | prec11 | 43.1 | 1.4 | prec1 | 15.7 | 2.5 | prec8 | 36.7 | 0 | prec11 | 47.3 | 4.9 | prec1 | 25.5 | 16.4 |
| srad4 | 12.2 | 6 | srad4 | 12.1 | 42.9 | alt | 14.8 | 2.1 | srad4 | 22.1 | 35.8 | alt | 15.5 | 2.6 | alt | 22.2 | 2.6 |
| tavg5 | 9.3 | 0 | alt | 11.9 | 5.6 | bio18 | 12.2 | 13.8 | alt | 14.9 | 7.7 | srad4 | 10.5 | 36.6 | bio15 | 14.8 | 12.4 |
| tavg4 | 7.6 | 26.7 | prec1 | 10.1 | 4.3 | srad4 | 11.4 | 41.1 | wind2 | 7.6 | 1.7 | tmin4 | 6.9 | 0 | bio18 | 11.3 | 3 |
| sh_r_s | 7.1 | 0 | wind2 | 7.2 | 0 | wind3 | 8.6 | 0.5 | prec3 | 2.3 | 1.5 | bio17 | 4.5 | 4.1 | srad8 | 7.7 | 0.3 |
| bio15 | 6.4 | 44.3 | bio9 | 4.9 | 0.3 | bio15 | 6.8 | 11.2 | ndvi_s | 2.1 | 0.3 | prec10 | 2.3 | 0.9 | prec11 | 3.3 | 2.7 |
| tmin5 | 5.3 | 2.4 | prec3 | 4.6 | 2.6 | prec7 | 5.3 | 0.7 | bio2 | 2 | 0.6 | tmin5 | 2.2 | 0.1 | srad4 | 2.3 | 23.2 |
| bio14 | 3.4 | 1.8 | ndvi_s | 1.3 | 0.6 | prec11 | 4.8 | 0.2 | srad9 | 1.9 | 3.6 | tavg4 | 1.3 | 0 | bio8 | 1.7 | 4.2 |
| sh_r_e | 2.9 | 0 | prec5 | 3.7 | 0.4 | bio1 | 1.1 | 0 | prec5 | 1.6 | 2.3 | ||||||
| srad6 | 2.9 | 0 | bio4 | 3.4 | 2.3 | ndvi_w | 1 | 0 | slope | 1.1 | 5.8 | ||||||
| bio1 | 2.6 | 0 | bio8 | 2.5 | 0.9 | tmax4 | 1 | 0 | |||||||||
| tmax4 | 1.6 | 0 | srad10 | 1.7 | 0.3 | ||||||||||||
| bio13 | 1.5 | 6.3 | wind6 | 1.5 | 2.3 | ||||||||||||
| bio17 | 1.5 | 0 | |||||||||||||||
| ndvi_s | 1.3 | 0 | |||||||||||||||
| prec11 | 1.2 | 4 | |||||||||||||||
| aspect | 1 | 0 | |||||||||||||||
По данным оценки численности населения в регионах (Численность…, 2019) плотность населения на Центральном Кавказе (Ингушетия, Северная Осетия, Кабардино-Балкария, Карачаево-Черкесия) составляет 81.7 чел./км2, а на Западном Кавказе (Краснодарский край, Адыгея) 66.5 чел./км2. Эти данные сопоставимы с таковыми для провинций Ирана, в которых отмечено присутствие леопарда (Sanei, Zakaria, 2011): средняя плотность на провинцию составляет 85.2 чел./км2, а медиальная – 62 чел./км2 (Provinces…, 2019). Таким образом, разница в плотности населения Западного и Центрального Кавказа, вероятно, не будет играть первостепенной роли при выборе леопардами участка обитания.
Из результатов анализа, основанных только на данных, поступивших от GPS ошейников, следует, что из 115 факторов интерполяционной модели, общей для всех особей, в построение объективной модели потенциально пригодных местообитаний вклад более 1% вносят только 10. Отдельно для самок количество определяющих факторов больше и составляет 14 факторов (табл. 3а). Для самца Ахуна их число еще больше, но это может быть связано с небольшим количеством точек, полученных от его ошейника (табл. 3б), а интерпретация этих данных по этой особи не может быть объективна. Среди основных факторов, в комплексе максимально формирующих модель потенциальных местообитаний леопарда, следует особенно отметить количество осадков в наиболее теплый квартал (в среднем около 300 мм) и в августе (порядка 100 мм); высоту над уровнем моря (от 500 до 2000 м над ур. м.); скорость ветра в марте (рис. 4).
Рис. 4.
Значения различных факторов, вносящих наибольший вклад в формирование потенциально пригодных местообитаний переднеазиатского леопарда на территории Кавказа: а – высота (м над ур. м.); б – количество осадков в августе; в – количество осадков в наиболее теплый квартал; г – средняя скорость ветра в марте.

Совокупный вклад этих четырех факторов в модель составляет более 60%. Индекс, отражающий глубину снежного покрова, не вносит значительный общий вклад в построение модели, однако обладает высоким коэффициентом пермутации. Если сравнивать общую модель и модель, рассчитанную для самок и самцов отдельно, то 5 факторов с наибольшим вкладом являются общими определяющими, наряду с высотой, осадками в наиболее теплый период и скоростью ветра, также уровень солнечной радиации в сентябре и количество осадков в январе (табл. 2).
При сравнении факторов, определяющих модели для каждой особи леопардов, выпущенных на Центральном Кавказе, на ведущие места выходит количество осадков в январе и сезонность осадков в целом, а для западно-кавказских особей высоко влияние осадков в ноябре и уровень солнечной радиации в апреле.
Существенного влияния факторов, характеризующих рельеф, не выявлено. В целом, наиболее значимыми, в различных комбинациях, являются климатические факторы и для отдельных особей вегетационный индекс. Территории с наиболее высокими значениями вероятности встречаемости локализуются в предгорьях и среднегорьях с травянистой растительностью, где при малой степени зависимости от экспозиции, преобладают средние значения глубины снежного покрова в зимний период (до 30 см).
Верификация модели
Одной из важных задач в ходе моделирования потенциально пригодных территорий является верификация модели. По результатам верификации на основе локаций от ошейника вероятность встречаемости самки леопарда в спрогнозированных для вида точках составляет вероятность 0.65, при медианном значении 0.66. Для моделей построенным по всем самкам эти значения сопоставимы (среднее 0.65, медиана 0.69) – табл. 4. Непосредственно по самке Волна значения 2019 года (модель с полным набором точек) характеризуется более высокими значениями вероятности встречаемости по сравнению с предварительной моделью.
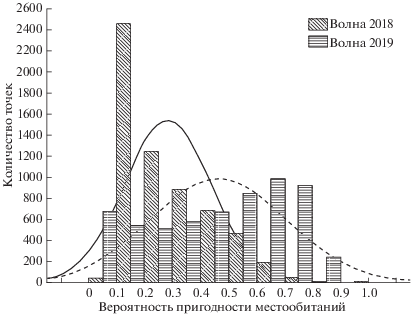
Подобные расхождения значений вероятности пригодности местообитаний для различных моделей, а также отсутствие значений вероятности пригодности местообитаний, близких к 1 даже в максимуме, можно объяснить как спецификой перемещения Волны (хотя и несущей черты вида в целом, но обладающей и индивидуальными характеристиками), так и малым отрезком времени, которое прошло с момента выпуска. Также это может объясняться тем, что участок обитания Волны еще не был сформирован. В пользу последнего говорит также и сравнительная гистограмма распределения значений вероятности встречаемости (пригодности местообитаний) для моделей 2018 и 2019 гг. (рис. 5). На рисунке видно, что распределение 2019 г. характеризуется как более равномерное, с увеличенной долей оптимальных участков, оно приближено к нормальному, и это, в свою очередь может свидетельствовать о прогрессе в формировании участка обитания (Hernandez-Blanco et al., 2020 in print).
Рис. 5.
Сравнительная гистограмма частот вероятности встречаемости (индекса пригодности местообитаний) для моделей Волны 2018 и 2019 гг.
Описанный выше подход нуждается в дальнейшей отработке и на других особях, однако показывает, на наш взгляд, возможность его предварительного применения для прогнозов использования пространства особями после выпуска, что облегчает оперативное реагирование в случае возникновения спонтанных ситуаций потенциального конфликта хищник–человек.
Модели, подобные полученным в данной работе, могут быть использованы для оценки участков, потенциально пригодных для вида, в том числе для корректировки географической локализации мест выпуска в последующем, проведения биотехнических мероприятий, оптимизации природоохранных мер и использования ресурсов и т.д. Они могут пригодиться для прогнозирования возможных путей перемещения животных и, соответственно рисков, сопряженных с возможным выходом животных к населенным пунктам и возникновению конфликтных ситуаций. Для иллюстрации подобных возможностей приведен результат анализа модели по полному треку Волны с наложением смоделированных территорий на административную карту Центрального Кавказа (рис. 6).
Рис. 6.
Модель потенциально пригодных местообитаний Волны, наложенная на административно-территориальную карту территории Кабардино-Балкарии, Северной и Южной Осетии.
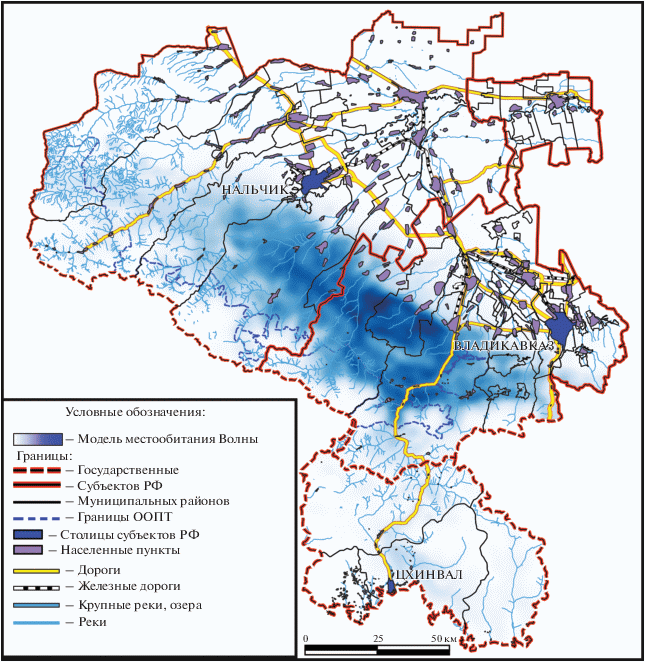
Согласно данной модели, основные участки потенциально пригодных местообитаний на Центральном Кавказе расположены в предгорной части Северной Осетии, частично затрагивая район Северо-Юрской депрессии Центрального Кавказа (южные и юго-западные отроги Скалистого Хребта). Административно эти участки охватывают территории Алагирского, Ирафского и Дигорского районов Республики Северная Осетия-Алания и Лескеского, Черекского и частично Чегемского районов Кабардино-Балкарии. Административные районы и населенные пункты, по которым вычислена высокая доля вероятности посещения леопардом по данным модели (от 0.7), характеризуются общими чертами, такими как невысокая численность и плотность населения, более низкий уровень разведения крупного и мелкого рогатого скота, выпасаемого на летних пастбищах в горной части (табл. 5), согласно данным о получаемой сельхозпродукции (Балаева, 2005; Хапачев, Боготов, 2008).
Таблица 5
Таблица 4. Накопительная верификация моделей по локациям самки по кличке Волна
| Модель | N (количество точек локаций Волны, включенных в анализ) | Значения вероятности пригодности местообитаний | ||||||
|---|---|---|---|---|---|---|---|---|
| среднее значение | медиана | минимум | максимум | стандартное отклонение | коэффициент вариации | ошибка среднего | ||
| Все леопарды | 3368 | 0.65 | 0.66 | 0.20 | 0.86 | 0.11 | 16.5 | 0.002 |
| Все самки | 3368 | 0.65 | 0.69 | 0.03 | 0.86 | 0.15 | 22.2 | 0.002 |
| Волна 2018 | 3368 | 0.47 | 0.54 | 0.02 | 0.69 | 0.19 | 39.7 | 0.003 |
| Волна 2019 | 3368 | 0.51 | 0.60 | 0.03 | 0.80 | 0.20 | 40.78 | 0.005 |
Таблица 6
Таблица 6. Численность населения в населенных пунктах анализируемой зоны
| № | Название (англ.) | Название (рус.) | Население, чел. | Вероятность встречаемости на основании моделей самки Волны | ||
|---|---|---|---|---|---|---|
| первичная модель | 2018 г. | 2019 г. | ||||
| Кабардино-Балкария | ||||||
| 1 | Verkhniy Lesken | Верхний Лескен | 146 | 0.28 | 0.52 | 0.61 |
| 2 | Verkhnya Balkariya | Верхняя Балкария | 4272 | 0.21 | 0.52 | 0.51 |
| 3 | Verkhnyaya Zhemtala | Верхняя Жемтала | 1638 | 0.21 | 0.55 | 0.61 |
| 4 | Karasu | Карасу | 517 | 0.15 | 0.59 | 0.71 |
| 5 | Tashli-Tala | Ташлы-Тала | 646 | 0.16 | 0.77 | 0.71 |
| Северная Осетия-Алания | ||||||
| 6 | Dzivgis | Дзивгис | 15 | 0.45 | 0.52 | 0.53 |
| 7 | Verkhny Fiagdon | Верхний Фиагдон | 1483 | 0.36 | 0.54 | 0.62 |
| 8 | Gorny Kartsa | Горный Картса | 46 | 0.47 | 0.54 | 0.37 |
| 9 | Verkhniy Biragzang | Верхний Бирагзанг | 860 | 0.52 | 0.56 | 0.53 |
| 10 | Khidikus | Хидикус | 117 | 0.46 | 0.57 | 0.74 |
| 11 | Verkhniy Zgid | Верхний Згид | 204 | 0.55 | 0.58 | 0.60 |
| 12 | Sadon | Садон | 492 | 0.52 | 0.58 | 0.59 |
| 13 | Mizur | Мизур | 3166 | 0.61 | 0.65 | 0.68 |
| 14 | Buron | Бурон | 548 | 0.73 | 0.69 | 0.57 |
| 15 | Akharisar | Ахарисар | 350 | 0.38 | 0.74 | 0.78 |
Сравнительный анализ первичной модели с прогнозами 2018 и 2019 гг. показал увеличение вероятности встречи самки Волна в окрестностях населенных пунктов Северной Осетии и Кабардино-Балкарии и смещение потенциально избираемых ею местообитаний на северо-запад от места выпуска (табл. 6, рис. 7). Впоследствии в ходе освоения территории она подходилa близко к ряду населенных пунктов в соответствии с прогнозами. Для модели 2018 г. информация подобного рода имеет особенное значение для определения приоритетных районов для просветительской работы с населением, а также локального повышения охранных мер. Тем не менее, за анализируемый период не было зафиксировано ни одной реальной конфликтной ситуации, созданной леопардом в отношении человека или принадлежащего ему скота, что говорит о полноценном поведении избегания человека выпущенным животным.
Таблица 7.
Таблица 5. Численность, плотность населения и поголовье крупного (КРС) и мелкого (МРС) рогатого скота по административным единицам с высокими значениями вероятности встречи самки Волна по данным модели (Поголовье …, 2019; Численность …, 2019)
| Республика, муниципальный район | Численность населения, чел. | Плотность населения, чел./км2 | Поголовье скота,тыс. голов | |
|---|---|---|---|---|
| КРС | МРС | |||
| Кабардино-Балкария | 866 219 | 69.5 | 265.1 | 375.9 |
| Лескенский район | 29 612 | 56.6 | 15.9 | 8.6 |
| Черекский район | 28 081 | 12.7 | 32.5 | 119.8 |
| Чегемский район | 69 149 | 46.0 | 29.5 | 32.0 |
| Северная Осетия-Алания | 699 253 | 87.55 | 91.2 | 74.8 |
| Алагирский район | 36 598 | 18.2 | Нет данных | Нет данных |
| Ирафский район | 15 107 | 11.2) | Нет данных | Нет данных |
| Дигорский район | 18 283 | 31.9 | Нет данных | Нет данных |
ЗАКЛЮЧЕНИЕ
Площадь оптимальных местообитаний для переднеазиатского леопарда в районе исследований составляет около 4500 км2 и может быть пригодна для использования леопардами в количестве от 10 до 30 особей, при том, что самцы смогут освоить большие по площади территории с более разнообразными природно-климатическими условиями относительно самок. Основными факторами, определяющими выбор леопардами участков обитания, являются количество осадков в наиболее теплый квартал, высота над уровнем моря (до 2000 м над ур. м.), крутизна склона (до 30°) и вегетационный индекс, характеризующий травянистые склоны. Также играет роль вариация глубины снежного покрова на склонах.
Хотя территория Западного Кавказа предоставляет более широкий спектр оптимальных для вида условий (более благоприятные ландшафтно-климатические и социально-экономические условия), она наиболее удалена от основных маршрутов расселения и миграции леопардов, которые также вычислили в результате проведенной работы. Верификация модели показала применимость методов пространственного анализа и моделирования для выявления и оценки потенциально пригодных местообитаний и мест перехода для особей вида, оценки риска возникновения возможных конфликтов и может быть использована для разработки научно-обоснованных природоохранных мероприятий, а также для определения приоритетных районов при планировании работы по экологическому просвещению населения. Также модель может быть применена для локального повышения охранных мер.
В будущем модель может быть улучшена с учетом влияния на формирование леопардами участков с учетом более точных данных антропогенного (присутствия инфраструктурных объектов на выбор леопардами участков обитания), а также таких биотических факторов, как распределение и плотность основных объектов питания и потенциальных конкурентов.
Список литературы
Балаева С.И. Состояние и перспективы развития молочного комплекса Кабардино-Балкарской республики // Региональная экономика: теория и практика. 2005. № 5. С. 55–57.
Верещагин Н.К. Млекопитающие Кавказа. Л.: “Ленинградское отделение Издательства Академии наук СССР”, 1959. 704 с.
Всероссийская перепись населения, 2010.
https://www.gks.ru/free_doc/new_site/perepis2010/croc/ perepis_itogi1612.htm
Гвоздецкий Н.А. Кавказ. М.: Географгиз, 1963. 262 с.
Гептнер В.Г., Слудский А.А. Млекопитающие Советского Союза. Т. 2. Ч. 2. Хищные (гиены и кошки). М.: Высшая школа, 1972. 552 с.
Герасимов И.П., Бавар И., Дреш Ж. Альпы-Кавказ: современные проблемы конструктивной географии горных стран. М.: Наука, 1980. 325 с.
Гольцман М.Е., Сушко Е.Д., Доронина Л.О., Крученкова Е.П. Индивидуум-ориентированная модель популяционной динамики песца острова Медный (Vulpes lagopus semenovi, Красная Книга РФ) // Принц. экол. 2016. Т. 5. № 3. С. 42.
Гольцман М.Е., Сушко Е.Д., Доронина Л.О., Крученкова Е.П. Индивидуум ориентированная модель популяционной динамики песца (Vulpes lagopus semenovi) на о-ве Медном (Командорские острова) // Зоол. журн. 2018. Т. 97. № 11. С. 1–18.
Добрынин Д.В., Рожнов В.В., Савельев А.А., Сухова О.В., Ячменникова А.А. Комплексирование данных мечения животных GPS-передатчиками и материалов мультиспектральной космической съемки для детальной характеристики местообитаний // Исслед. Земли из космоса. 2017. № 3. С. 40–52.
Жашуев А.Ж., Пшегусов Р.Х. Эколого-географический анализ избранных видов печеночников природных и антропогенных экосистем зоны рекреации на Центральном Кавказе) // Изв. Уфимского научного центра РАН. 2016. № 4. С. 56–61. О. В.
Жу Ш., Чю И., Лю И., Добрынин Д.В., Сухова О.В., Котлов И.П., Сандлерский Р.Б., Ячменникова А.А., Рожнов В.В. Структура потенциальных местообитаний амурского тигра (Panthera tigris altaica) и оценка его кормовой базы на территории национального парка Тайпингоу (Китай) по данным дистанционного зондирования Земли // Исслед. Земли из космоса. 2019. № 4. С. 60–86.
Красная книга Российской Федерации (животные) / Под ред. РАН; гл. редкол.: В.И. Данилов-Данильян и др. М.: АСТ: Астрель, 2001. 862 с. ISBN 5-17-005792-X, 5-271-00651-4.
Огурцов С.С. Моделирование пригодности местообитаний и распределения бурого медведя (Ursus arctos) в подзоне южной тайги с помощью метода максимальной энтропии // Nature Conservation Research. Заповедная наука. 2019. Т. 4. № 4. С. 34–64.https://doi.org/10.24189/ncr.2019.061
Особо охраняемые природные территории России: современное состояние и перспективы развития, авторы-составители В.Г. Кревер, М.С. Стишов, И.А. Онуфреня. Москва: WWF России, 2009. 456 с.
Поголовье скота в Российской Федерации в 2018 году. Электронный ресурс: https://www.gks.ru/storage/mediabank/pog-skot_18(1).rar. Дата обращения: 5 декабря 2019 г.
Программа по восстановлению (реинтродукции) переднеазиатского леопарда на Кавказе / Под ред. Рожнова В.В., Лукаревского В.С. Москва: КМК – ИПЭЭ РАН, 2008. 65 с.
Пшегусов Р.Х. Технологии мониторинга популяций редких видов: теория и практика применения в условиях горных экосистем // Материалы Международной конференции “Хищные птицы Северного Кавказа и сопредельных регионов: Распространение, экология, динамика популяций, охрана”. Сочи, 8–10 апреля 2014 г. Ростов-на-Дону: Издательство Южного федерального университета, 2014. С. 42–53.
Пшегусов Р.Х., Ханов З.М. Технологии дистанционного мониторинга: методы прогнозирования и оценки местообитаний редких видов лишайников на Центральном Кавказе” // Тезисы докладов III (XI) Международной Ботанической конференции молодых ученых в Санкт-Петербурге. 4–9 октября 2015. С. 39.
Родникова А.Ю., Каримова Т.Ю., Лущекина А.А., Ячменникова А.А., Грибов К.А., Рожнов В.В., Гольцман М.Е. Индивидуум-ориентированная модель популяционной динамики сайгака Северо-Западного Прикаспия // Saiga News. 2018. Вып. 24. С. 23–26.
Рожнов В.В., Ячменникова А.А., Найденко С.В., Эрнандес-Бланко Х.А., Чистополова М.Д., Сорокин П.А., Добрынин Д.В., Сухова О.В., Поярков А.Д., Дронова Н.А., Трепет С.А., Пхитиков А.Б., Пшегусов Р.Х., Магомедов М.Р.Д. Мониторинг переднеазиатского леопарда и других крупных кошек. М.: КМК, 2018. 121 с.
Рожнов В.В., Чистополова М.Д., Ячменникова А.А., Эрнандес-Бланко Х.А., Сорокин П.А., Найденко С.В. 2019. Ориентация крупных хищных млекопитающих после выпуска в природу на примере тигра и леопарда // Тезисы конференции “Ориентация и навигация животных”. 2–4 октября 2019 г. Москва. М.: ИПЭЭ РАН, КМК. С. 65.
Семёнов У.А. Центр восстановления леопарда на Кавказе – стратегия реинтродукции // Тр. Сочинского национального парка. 2016. Вып. 10. М.: КМК. 112 с.
Хапачев А.Б., Боготов Х.Л. Состояние и направления развития аграрного сектора Кабардино-Балкарской республики // Региональная экономика: теория и практика. 2008. № 31. С. 66–75.
Цепкова Н.Л., Пшегусов Р.Х., Ханов З.М., Жашуев А.Ж. Оценка распространения травяных сообществ на основе данных дистанционного зондирования в мониторинге состояния горных лугов Центрального Кавказа (Кабардино-Балкария) // Изв. Самарского научного центра Российской академии наук. 2015. Т. 17. № 4–2. С. 428–432.
Численность населения Российской Федерации по муниципальным образованиям на 1 января 2019 года. Электронный ресурс: http://www.gks.ru/free_doc/ doc_2019/bul_dr/mun_obr2019.rar. Дата обращения 31 июля 2019.
Чистополова М.Д., Рожнов В.В., Эрнандес-Бланко Х.А., Найденко С.В., Сорокин П.А. Новый аналитический подход к изучению пространственной структуры популяции дальневосточного леопарда (Panthera pardus orientalis) // Экол. 2018. № 6. С. 448–458.
Araujo M.B., Peterson A.T. Uses and misuses of bioclimatic envelope modeling // Ecol. 2012. V. 93. P. 1527–1539. https://doi.org/10.1890/11-1930.1
Austin M.P. Spatial prediction of species distribution: an interface between ecological theory and statistical modelling // Ecol. Model. 2002. V. 157. P. 101–118.
Baldwin R.A. Use of maximum entropy modeling in wildlife research. Entropy. 2009. Vol. 11. Iss. 4. P. 854-866.
Bleyhl B., Sipko T., Bragina E., Heidelberg A., Leitão P. J., Pshegusov R., Radeloff V.C., Trepet S., Weinberg P., Kuemmerle T. Potential European bison habitat in the Caucasus // Conference program and abstracts “International Biogeography Society 7th Biennial Meeting”. 8–12 January 2015, Bayreuth, Germany. Frontiers of Biogeography. V. 6. Suppl. 1. International Biogeography Society. P. 143.
Buckley A. Understanding curvature rasters [Electronic resource]. 2010. https://blogs.esri. com/esri/arcgis/2010/10/ 27/understanding-curvature-rasters. Request: 5.12.2018.
Carter N., Levin S., Barlow A., Grimm V. Modeling tiger population and territory dynamics using an agent-based approach // Ecol. Model. 2015. № 312. P. 347–362.
CITES. Конвенция о международной торговле видами дикой фауны и флоры, находящимися под угрозой исчезновения, Принята 3 марта 1973 года https://www.un.org/ ru/documents/decl_conv/conventions/cites.shtml; https:// cites.org/eng/app/appendices.php
De Marco P., Nobrega C.C. Evaluating collinearity effects on species distribution models: An approach based on virtual species simulation. 2018. PLoS ONE V. 13 № 9: e0202403. https://doi.org/10.1371/journal.pone.0202403
Dormann C.F., Purschke O., Marquez J.R., Lautenbach S., Schroeder B., Garcia M.JR. Components of uncertainty in species distribution analysis: a case study of the Great Grey Shrike // Ecol. 2008. V. 89. P. 3371–3386. https://doi.org/10.1890/07-1772.1
Elith J., Graham C.H., Anderson R.P., Dudík M., Ferrier S., Guisan A., Hijmans R.J., Huettmann F., Leathwick J.R., Lehmann A., Li J., Lohmann L.G., Loiselle B.A., Manion G., Moritz C., Nakamura M., Nakazawa Y., Overton J.Mc.M, Peterson A.T., Phillips S.J., Richardson K., Scachetti-Pereira R., Schapire R.E., Soberón J., Williams S., Wisz M.S., Zimmermann N.E. Novel methods improve prediction of species’ distributions from occurrence data // Ecography. 2006. V. 29. I. 2. P. 129–151.
Elith J., Hastie T., Dudík M., Chee Yu.E., Yates C. A statistical explanation of MAXENT for ecologists // Diversity and Distributions. 2010. V. 17. P. 43–57.https://doi.org/10.1111/j.1472-4642.2010.00725.x
Farhadinia M.S., Johnson P.J., Macdonald D.W., Hunter L.T.B. Anchoring and adjusting amidst humans: Ranging behavior of Persian leopards along the Iran-Turkmenistan borderland // PLoS ONE. V. 13. № 5. e0196602. 2018. https://doi.org/10.1371/journal.pone.0196602
Giovanelli J., Siqueira M., Haddad C., Alexandrino J. Modeling a spatially restricted distribution in the Neotropics: How the size of calibration area affects the performance of five presence-only methods // Ecol. Model. 2010. V. 221. P. 215–224. https://doi.org/10.1016/j.ecolmodel.2009.10.009
Guisan A., Graham C., Elith J., Huettmann F., Dudik M., Ferrier S., Hijmans R., Lehmann A., Li J., Lohmann L., Loi-selle B., Manion G., Moritz C., Nakamura M., Nakazawa Yo., Overton J., Peterson A., Phillips S., Richardson K., Zimmermann N. Sensitivity of predictive species distribution models to change in grain size // Diversity and Distributions. V. 13. I. 3. 2007. P. 332–340.https://doi.org/10.1111/j.1472-4642.2007.00342.x
Guisan A., Zimmermann N., Guisan A., Zimmermann N.E. Predictive habitat distribution models in ecology // Ecol. Model. 2000. V. 135. P. 147–186.https://doi.org/10.1016/S0304-3800(00)00354-9
Guo Q., Liu Y. ModEco: an integrated software package for ecological niche modeling // Ecography. 2010. V. 33. P. 1–6.
Hayward M.W., Somers M.J. Reintroduction of Top-Order Predators: Conservation Science and Practice Series. Oxford, Blackwell Publishing Ltd., 2009. 449 p.
Hebblewhite M., Miquelle D.G., Robinson H., Pikunovd D.G., Dunishenko Y.M., Aramilev V.V., Nikolaev I.G., Salkina G.P., Seryodkin I.V., Gaponov V.V., Litvinov M.N., Kostyria A.V., Fomenko P.V., Murzin A.A. Including biotic interactions with ungulate prey and humans improves habitat conservation modeling for endangered Amur tigers in the Russian Far East // Biol. Conserv. V. 178. 2014. P. 50–64.
Hernandez-Blanco J.A., Chistopolova M.D., Yachmennikova A.A., Trepet S.A., Pkhitikov A.B., Naidenko S.V., Sorokin P.A., Dronova N.A., Rozhnov V.V. Space use and Dynamic Interaction Patterns of Persian Leopards (Panthera pardus saxicolor) after reintroduction into Western Caucasus // PLoS One, 2020, in print.
Hijmans R.J., Cameron S.E., Parra J.L., Jones P.G., Jarvis A. Very high resolution interpolated climate surfaces for global land areas // Intern. Journ. of Climatol. 2005. V. 25. P. 1965–1978.
Jiang G., Oi J., Gu J., Chang Y., Shi Q., Lui P. 2016. Population and habitat of Amur leopard in China / Eds. by Jiang G. Science Press Beijing, 2016. 202 p.
Kanagaraj R., Wiegand T., Kramer-Schadt S., Goyal S.P. Using individual-based movement models to assess inter-patch connectivity for large carnivores in fragmented landscapes // Biol. Conserv. 2013. V. 167. P. 298–309.
Koshkarev E.P. Characteristics of snow leopard (Uncia uncia) movements in the Tien Shan // International Pedigree Book of Snow Leopards. 1984. V. 4. P. 15–21.
Krever V., Zazanashvili N., Jungius H., Petelin D. Biodiversity of the Caucasus: An Analysis of Biodiversity and Current Threats and Initial Investment Portfolio. 2001.
McNally A., Arsenault K., Kumar S., Shukla S., Peterson P., Wang S., Funk C., Peters-Lidard C.D., Verdin J.P. A land data assimilation system for sub-Saharan Africa food and water security applications // Sci. Data. 2017. № 4. Art. No: 170012. https://doi.org/10.1038/sdata.2017.12
Moore I.D., Grayson R.B., Ladson A.R. Digital terrain modelling: a review of hydrological, geomorphological, and biological applications // Hydrological processes. Special Issue: Digital Terrain Modelling in Hydrology. 1991. V. 5. I. 1. P. 3–30.
Naqibzadeh A., Rezaei N., Sarhangzadeh J., Sayedi N. Desirability Modeling Wild Sheep Habitat in Wildlife Refuge of Burueiyeh in Yazd Province by Using Maximum Entropy Method (MaxEnt) // Indo-Iranian Journ. 2019. V. 10. № 4. P. 75–82.
Phillips S.J., Anderson R.P., Schapire R.E. Maximum entropy modeling of species geographic distribution // Ecol. Model. 2006. V. 190. I. 3–4. P. 231–259. https://doi.org/10.1016/j.ecolmodel.2005.03.026
Phillips S., Dudík M. Modeling of species distributions with MAXENT: new extensions and a comprehensive evaluation // Ecography. 2008. V. 31. P. 161–175. https://doi.org/10.1111/j.0906-7590.2008.5203.x
Provinces of Iran. Электронный ресурс: https://en.wikipedia.org/wiki/Provinces_of_Iran Дата обращения: 5 декабря 2019 г.
Rondinini C., Stuart S., Boitani L. Habitat Suitability Models and the Shortfall in Conservation Planning for African Vertebrates // Conserv. Biol. 2005. V. 19. № 5. P. 1488–1497.
Rozhnov V.V., Yachmennikova A.A., Hernandez-Blanco J.A., Naidenko S.V., Chistopolova M.D., Sorokin P.A., Dobrynin D.V., Sukhova O.V., Poyarkov A.D., Dronova N.A., Trepet S.A., Pkhitikov A.B., Pshegusov R.H., Magomedov M.-R.D. Study and Monitoring of Big Cats in Russia. Moscow: IEE RAS, KMK, 2019. 120 p.
Safford R.K. Modeling critical winter habitat of four ungulate species in the Robson Walley, British Columbia // J. of Ecosystems and Management. 2004. V. 4. № 2. P. 1–13.
Sanei A., Zakaria M. Survival of the Persian leopard (Panthera pardus saxicolor) in Iran: primary threats and human-leopard conflicts // Asia Life Sciences Supplement. 2011. V. 7. P. 31–39.
Sarrazin F., Barbault R. Reintroduction: Challenges and Lessons for Basic Ecology // Trends in Ecol. & Evol. 1996. V. 11. P. 474–478. https://doi.org/10.1016/0169-5347(96)20092-8
Stigall A.L. Using ecological niche modeling to evaluate niche stability in deep time // J. of Biogeogr. 2012. V. 39. P. 772–781. https://doi.org/10.1111/j.1365- 2699.2011.02651.x
Stockwell D., Peters D. The GARP modeling system: Problems and solutions to automated spatial prediction // Intern. J. of Geogr. Inform. Sc. 1999. V. 13. P. 143–158. https://doi.org/10.1080/136588199241391
Studley H., Weber K.T. Comparison of Image Resampling Techniques for Satellite Imagery / Eds. by Weber K.T. and Davis K. Final Report: Assessing Post-Fire Recovery of Sagebrush-Steppe Rangelands in Southeastern Idaho. 2011. P. 185–196. (https://pdfs.semanticscholar.org/fa31/ 6fc18b22fe466f818bcea0edae76d08c91f9.pdf)
Swanepoel L.H., van Hoven W. Home range and movement of leopard (Panthera pardus) on selected game and livestock ranches in the Waterberg, Limpopo South Africa // Ecology and conservation of leopards, Panthera pardus, on selected game ranches in the Waterberg region, Limpopo, South Africa. Univ. of Pretoria, 2008. P. 23–47.
West A.M., Kumar S., Brown C.S., Stohlgren T.J., Bromberg J. Field validation of an invasive species Maxent model // Ecol. Inform. 2016. Vol. 36. P. 126–134.https://doi.org/10.1016/j.ecoinf.2016.11.001
Zevenbergen L.W., Thorne C.R. Quantitative analysis of land surface topography // Earth surface proc. and landforms. 1987. V. 12. № 1. P. 12–56.
Дополнительные материалы отсутствуют.
Инструменты
Исследование Земли из Космоса