

Экология, 2020, № 1, стр. 51-61
Влияние климатических факторов на эмиссию СО2 из почв в среднетаежных лесах Центральной Сибири: эмиссия как функция температуры и влажности почвы
А. В. Махныкина a, b, *, А. С. Прокушкин a, b, О. В. Меняйло b, С. В. Верховец a, И. И. Тычков a, А. В. Урбан b, А. В. Рубцов a, Н. Н. Кошурникова a, Е. А. Ваганов a, b
a Сибирский федеральный университет
660041 Красноярск, просп. Свободный, 79, Россия
b Институт леса им. В.Н. Сукачева СО РАН
660036 Красноярск, Академгородок, 50/28, Россия
* E-mail: amakhnykina@sfu-kras.ru
Поступила в редакцию 29.03.2019
После доработки 16.05.2019
Принята к публикации 18.06.2019
Аннотация
Почвенная эмиссия СО2 – один из важнейших компонентов глобального цикла углерода. В данной работе анализируется сезонная динамика почвенной эмиссии в разных типах земного покрова в подзоне средней тайги Центральной Сибири в течение пяти вегетационных сезонов. Показано, что для обширной территории сосновых лесов и их производных, сформированных на песчаных почвах, величина сезонной эмиссии СО2 определяется прежде всего условиями увлажнения, и лишь во вторую очередь температурным режимом и типом экосистемы. Роль типа леса проявляется при наиболее благоприятных по увлажнению условиях. Предложен подход по разделению сезона на сухой и влажный периоды в зависимости от пороговой влажности для участков с разными типами растительного покрова.
Взаимодействие климатических изменений и наземного цикла углерода – один из центральных вопросов современной экологии и биогеохимии. Эмиссия СО2 из почв в атмосферу – результат автотрофного корневого и гетеротрофного микробного дыхания [1, 2]. Временная динамика эмиссии СО2 обусловлена фенологическими особенностями растений, концентрацией в почвах лабильного углерода и азота [3], сообществом микроорганизмов и климатическими факторами территории, наиболее важными из которых являются температура и влажность почвы [4–6]. Параметризация взаимодействия между эмиссией СО2 и климатическими факторами, контролирующих этот процесс, имеет решающее значение для моделирования баланса углерода и понимания реакции биогеоценозов на изменение климата [7–9].
Повышение эмиссии СО2 при потеплении – ключевой механизм положительной обратной связи между экосистемами и глобальными изменениями климата [10–13]. Параметризация зависимости потоков СО2 из почвы от температуры является основным подходом в моделировании баланса углерода экосистем [5, 14–16], описываемая, как правило, через экспоненциальные функции отклика [9, 17]. Показано, что для биогеоценозов бореальной и умеренной зон почвенная эмиссия СО2 может описываться исключительно температурой [18, 19], тогда как в экосистемах с дефицитом осадков необходимо также учитывать влажность почв [20]. В настоящее время возрастание роли влагообеспеченности в формировании эмиссионных потоков в бореальных и умеренных лесах происходит из-за увеличения частоты и продолжительности засушливых периодов [21, 22].
Цель настоящей работы – установление влияния климатических факторов на поток СО2 из почв (на основе его межгодовой изменчивости) в нескольких типах лесных биогеоценозов подзоны средней тайги Центральной Сибири и оценка зависимости почвенной эмиссии СО2 от температуры и влажности почвы. Мы исходили из предположения, что зависимость потока СО2 от температуры неодинакова для разных типов леса и изменяется в течение вегетационного сезона. В задачи исследования входило: 1) оценить влияние гидротермических условий вегетационного сезона на формирование потока СО2; 2) охарактеризовать временные изменения почвенной эмиссии СО2 на участках с разными типами растительного покрова и 3) оценить зависимости почвенной эмиссии от температуры и влажности почвы в течение сезона. В бореальной зоне Сибири существуют обширные территории сосновых боров и их производных, произрастающих на песчаных почвах, где величина сезонной эмиссии СО2 определяется условиями увлажнения вегетационного сезона. Нами предложен новый подход к моделированию эмиссии СО2 в таких экосистемах путем выделения в вегетационном сезоне сухих и влажных периодов.
МАТЕРИАЛ И МЕТОДЫ
Характеристика климата и метеорологические условия в период исследования
Исследования проводили в среднетаежной подзоне Центральной Сибири, в южной части Туруханского района Красноярского края (60°26′ с.ш., 89°24′ в.д.). Климат региона резко-континентальный, среднегодовая температура воздуха составляет –3.5°С (1936–2017 гг.). Сумма температур выше 10°C варьирует в пределах 1200–1400°C. Самым теплым месяцем года является июль – среднемесячная температура составляет 18.1°C, среднемесячная температура самого холодного месяца (января) –23.8°C. Амплитуда колебания среднемесячных температур может достигать 42°C [23]. Среднемноголетнее среднее количество осадков за вегетационный сезон (июнь–сентябрь) для данной территории составляет 261 мм, а для всего года 594 мм (измерения с 1966 г.). Таким образом, на дождевые осадки за вегетационный сезон приходится около 44%.
Следует отметить, что за период метеонаблюдений с 1936 г. по 2017 г. в районе исследования (метеостанция Бор, источник – http://www.meteo.ru) среднегодовая температура воздуха увеличилась на 3.2°С. Самым теплым был 2015 г., когда среднегодовая температура составила –0.5°С. При этом температура воздуха пяти рассматриваемых вегетационных сезонов (2012, 2013, 2015–2017 гг.) (рис. 1) также превышала среднемноголетнее значение (13.2°С) на 0.7–3.4°С: в 2012 г. средняя температура воздуха составила 16.5 ± 0.5°С, в 2013 г. – 14.2 ± 0.6°С, в 2015 г. – 14.2 ± 0.6°С, в 2016 г. – 16.6 ± ± 0.5°С и в 2017 г. – 13.9 ± 0.6°С.
Рис. 1.
Средняя температура (1) и суммарное количество осадков (2) за сезон измерений (июнь–сентябрь) для 5-летнего периода и среднемноголетние значения этих параметров по метеостанции Бор (1936–2017 гг. – для температуры воздуха и 1966–2017 гг. – для количества осадков). Данные по температуре воздуха приведены со стандартными ошибками.

Отличительной чертой последнего десятилетия стала высокая вариабельность годового количества осадков, которая между отдельными годами достигала более 350 мм: например, сумма осадков за 2016 г. составила 385 мм, а за 2017 г. – 743 мм. При этом наиболее выраженная изменчивость поступления атмосферных осадков отмечена именно в течение вегетационного сезона. Так, значительный дефицит осадков относительно среднемноголетней нормы наблюдался в 2012 г. (54% от нормы) и 2013 г. (65%). Повышенное количество осадков отмечено в сезон 2015 г. – 126% от среднего многолетнего. Сезон 2017 г. можно охарактеризовать как наиболее близкий к средним значениям по количеству осадков (291 и 261 мм соответственно) и температуре воздуха (13.9 и 11.6°С) (см. рис. 1).
Объекты исследования
В пределах геоморфологически однородной поверхности (вершина увала) на расстоянии менее 1.5 км друг от друга были выбраны четыре экспериментальных участка (ЭУ) с различными типами земного покрова: сосняк лишайниковый, сосняк зеленомошный, смешанный лес и открытая антропогенно-нарушенная почва без растительности. Описание экосистем приведено в табл. 1.
Таблица 1.
Описание экосистем, в которых проводились измерения (приведены средние значения и стандартные отклонения (SD) за пять сезонов измерений)
| Характеристика | Сосняк лишайниковый | Сосняк зеленомошный | Смешанный лес | Нарушенный участок |
|---|---|---|---|---|
| Координаты | 60°47′57.3′′ с.ш., 89°21′22.7′′ в.д. |
60°48′00.8′′ с.ш., 89°21′04.7′′ в.д. |
60°47′52.2′′ с.ш., 89°21′13.1′′ в.д. |
60°47′57.4′′ с.ш., 89°21′01.7′′ в.д. |
| Температура почвы, °С | 13.81 ± 3.74 | 11.47 ± 2.42 | 12.98 ± 2.55 | 20.04 ± 7.23 |
| Влажность почвы, м3 м–3 | 0.21 ± 0.09 | 0.26 ± 0.12 | 0.25 ± 0.10 | 0.31 ± 0.09 |
| Содержание С, г С кг–1 почвы (до глубины 50 см) | 13.0 ± 1.5 | 7.0 ± 3.6 | 14.9 ± 1.4 | 7.3 |
| Доля корней, % от общего запаса Сорг (до глубины 50 см) | 42 | 48 | 41 | – |
| Содержание N, г N кг–1 почвы (до глубины 50 см) | 0.6 ± 0.1 | 0.3 ± 0.1 | 0.6 ± 0.2 | 0.4 |
| Возраст древостоя, лет | 73 | 119 | 27 | – |
| Преобладающие древесные породы | Pinus sylvestris L. | Pinus sylvestris L. | Pinus sylvestris L., Betula pendula Roth, Populus tremula L. | – |
| Доминанты живого напочвенного покрова | Cladonia stellaris (Opiz) Pouzar et Vezda, Cl. arbuscula (Wallr) Flot | Pleurozium schreberi (Brid.) Mitt, Dicranum polysetum Michx. | Cladonia stellaris (Opiz) Pouzar et Vezda, Cl. arbuscula (Wallr) Flot, Pleurozium schreberi (Brid.) Mitt, Dicranum polysetum Michx | – |
Почвы района исследований сформированы на гляциофлювиальных отложениях и характеризуются преобладанием песка в верхней части профиля. Как правило, ниже глубины 1 м отмечаются глинистые горизонты (линзы). На участках с ненарушенным растительным покровом почвы относятся к подзолам иллювиально-железистым с различным по мощности органическим горизонтом (маломощным в сосняке лишайниковом и смешанном лесу). По Международной классификации (WBR) почвы ЭУ относятся к подзолам (Podzols). Нарушенный участок характеризуется отсутствием верхних генетических горизонтов почвы. Запасы углерода в почве лесных биогеоценозов относительно невелики, составляя по нашим оценкам около 4 кг С м–2 в 2-метровой толще. На органический горизонт почвы приходится более 30% запасов всего почвенного органического вещества (ОВ) [24]. Запасы корневой фитомассы варьирует от 30 до 60% почвенного ОВ, содержание детрита составляет порядка 10%.
Измерение почвенной эмиссии СО2, температуры и влажности почв
Измерения почвенной эмиссии осуществляли с июня по сентябрь в течение пяти лет: 2012, 2013, 2015, 2016 и 2017 гг. (с середины сезона). На каждом ЭУ весной 2012 г. были установлены поливинилхлоридные (ПВХ) кольца диаметром 20 см, расположенные на расстоянии 1–1.5 м друг от друга: на лесопокрытых участках – по пять колец, на нарушенном – три кольца. Измерения потоков СО2 с поверхности почвы производили при помощи инфракрасного газоанализатора LI 8100A (Li-cor Inc., США) в дневное время в период с 11:00 до 16:00. Измерения осуществляли в трех повторностях, на основании которых в дальнейшем рассчитывали среднее значение, время измерения – 2 мин с промежутком между измерениями 30 сек. Детальное описание методики приведено нами в работе [25]. Количество измерений за сезон по всем участкам варьировало от 33 (сезон 2017 г.) до 189 (сезон 2013 г.). Общее количество измерений для участков за все сезоны составило: для сосняка лишайникового – 144, для сосняка зеленомошного – 139, для смешанного леса – 140, для нарушенного участка – 87 (три сезона измерений).
При каждом измерении потоков СО2 производили замеры температуры почвы (на глубине 5, 10 и 15 см) с помощью почвенного температурного датчика Soil Temperature Probe Type E (Omega, США) и объемной влажности почвы SWC (на глубине 5 см), используя влагомер Theta Probe Model ML (Delta T Devices Ltd., Великобритания).
Оценка зависимости эмисионных потоков СО2 от гидротермических параметров и выделение периодов засух
Анализ зависимости включал рассмотрение трех ключевых компонентов – почвенной эмиссии СО2, температуры и влажности почвы, соответствующих каждому измерению эмиссии. Величины почвенной эмиссии СО2 и температуры почвы за пять сезонов измерений были расположены в ряду увеличения влажности почвы для каждой экосистемы отдельно. На основе полученной выборки была построена экспоненциальная зависимость эмиссии от температуры почвы. Затем пошагово уменьшали объем выборки в рядах эмиссии и температуры почвы – минимальный объем выборки составил по три значения. Путем поочередного исключения значений от большей влажности к меньшей (т.е. выборка последовательно характеризовалась возрастающим дефицитом почвенной влаги) оценивали изменение коэффициента детерминации R2 между величинами. По критерию Фишера (F) был рассчитан коэффициент корреляции (rфакт) для зависимости почвенной эмиссии СО2 от температуры почвы при разном объеме выборки. Для всех рассмотренных объемов выборки (N – количество измерений в экосистеме) были определены значения статистически значимого коэффициента корреляции (rзнач) при P < 0.05.
Процедура определения пороговой влажности состояла в установлении конкретного минимального значения rфакт, при котором соблюдалось неравенство: rфакт – rзнач > 0 (рис. 2). Выделенные группы измерений рассматривали по разнице потоков СО2 для сухого (до пороговой влажности) и влажного (выше пороговой влажности) периодов. Изучение скорости потоков СО2 без лимитирующих по влажности условий позволило оценить температурную чувствительность каждого из типов биогеоценозов.
Рис. 2.
Зависимость коэффициентов корреляции (r) между почвенной эмиссией СО2 и температурой почвы от влажности почв и объема выборки для участков с разными типами растительного покрова: а – сосняк лишайниковый, б – сосняк зеленомошный, в – смешанный лес, г – нарушенный участок; rзнач – распределение значимого коэффициента корреляции между эмиссией и температурой почвы, rфакт – измеренные значения.

Расчет Q10 для влажного и сухого периодов
Коэффициент температурной чувствительности Q10 был рассчитан по уравнению Вант-Гоффа [26]:
где T1 и T2 – температура почвы, R1 и R2 – поток СО2 из почвы при температуре T1 и T2. Построена экспоненциальная зависимость эмиссии СО2 от температуры почв для двух групп – сухого и влажного периодов и для каждой экосистемы отдельно. По полученным уравнениям находили значения потоков (R1 и R2) для температур 5 и 15°C. Поскольку различия между T2 и T1 составляют ровно 10°C, коэффициент Q10 рассчитывали как соотношение величин потоков при 5 и 15°C.Статистический анализ данных
При оценке сезонной динамики почвенной эмиссии СО2 использовали трехфакторный дисперсионный анализ с повторяющимися измерениями: тип экосистемы, год и период сезона (на основе юлианского дня, DOY – день года). Кроме того, двухфакторный дисперсионный анализ использовался для оценки совокупного влияния климатических условий (температуры и влажности почвы) на сезонный поток СО2. После 3- и 2-факторного дисперсионного анализа было проведено попарное сравнение величин (post hoc) методом наименьших значимых различий (LSD).
Коэффициент температурной чувствительности (Q10) был получен для каждой экосистемы без повторностей, но для сухого и влажного периодов отдельно. Это дало возможность рассчитать значимость различий в Q10 для сухого и влажного периодов (тест Стьюдента), где экосистемы рассматривались как повторности.
РЕЗУЛЬТАТЫ И ИХ ОБСУЖДЕНИЕ
Гидротермический режим почв
Сход снежного покрова происходит в период с 4 (2012 г.) по 14 (2015, 2016 гг.) мая. Соответственно переход температур почвы к положительным значениям приходится, как правило, на первую декаду мая. Максимальные температуры почв на всех ЭУ наблюдаются в 3-й декаде июля. По температуре почв (табл. 2) участки располагаются в следующем порядке: нарушенный участок > сосняк лишайниковый > смешанный лес > сосняк зеленомошный (P < 0.05).
Таблица 2.
Температура (Ts, °C) и влажность почв (SWC, м3 м–3) для всех ЭУ (приведены средние значения и стандартные ошибки (SE))
| Месяц | Сосняк лишайниковый | Сосняк зеленомошный | Смешанный лес | Нарушенный участок | ||||
|---|---|---|---|---|---|---|---|---|
| Ts | SWC | Ts | SWC | Ts | SWC | Ts | SWC | |
| 2012 г. | ||||||||
| Июнь | 16.5 ± 1.2 | 0.11 ± 0.01 | 10.4 ± 1.6 | 0.11 ± 0.03 | 13.3 ± 1.0 | 0.17 ± 0.03 | 31.1 ± 0.9 | 0.21 ± 0.01 |
| Июль | 18.3 ± 1.1 | 0.10 ± 0.01 | 13.2 ± 1.7 | 0.07 ± 0.01 | 15.9 ± 1.0 | 0.11 ± 0.01 | 22.3 ± 0.2 | 0.17 ± 0.01 |
| Август | 11.9 ± 0.3 | 0.22 ± 0.02 | 9.7 ± 0.1 | 0.20 ± 0.03 | 11.5 ± 0.2 | 0.21 ± 0.02 | 16.9 ± 0.1 | 0.20 ± 0.01 |
| Сентябрь | 10.2 ± 0.2 | 0.34 ± 0.20 | 9.7 ± 0.4 | 0.29 ± 0.04 | 10.5 ± 0.4 | 0.37 ± 0.05 | Нет данных | Нет данных |
| 2013 г. | ||||||||
| Июнь | 13.5 ± 0.3 | 0.23 ± 0.01 | 8.6 ± 0.2 | 0.30 ± 0.01 | 11.1 ± 0.2 | 0.32 ± 0.01 | 22.3 ± 0.3 | 0.42 ± 0.01 |
| Июль | 15.0 ± 0.3 | 0.18 ± 0.01 | 11.4 ± 0.2 | 0.23 ± 0.01 | 13.4 ± 0.1 | 0.24 ± 0.01 | 25.9 ± 0.2 | 0.28 ± 0.01 |
| Август | 15.2 ± 0.2 | 0.24 ± 0.01 | 12.7 ± 0.1 | 0.24 ± 0.01 | 14.2 ± 0.1 | 0.24 ± 0.01 | 20.7 ± 0.2 | 0.27 ± 0.01 |
| Сентябрь | 8.2 ± 0.1 | 0.28 ± 0.01 | 8.9 ± 0.04 | 0.41 ± 0.02 | 8.9 ± 0.04 | 0.37 ± 0.01 | 8.9 ± 0.1 | 0.39 ± 0.01 |
| 2015 г. | ||||||||
| Июнь | 15.2 ± 0.7 | 0.18 ± 0.01 | 12.1 ± 0.7 | 0.27 ± 0.02 | 13.6 ± 0.5 | 0.24 ± 0.02 | 24.5 ± 0.1 | 0.39 ± 0.01 |
| Июль | 17.1 ± 0.4 | 0.23 ± 0.01 | 14.9 ± 0.5 | 0.24 ± 0.02 | 15.9 ± 0.4 | 0.24 ± 0.02 | 25.4 ± 0.2 | 0.27 ± 0.01 |
| Август | 14.4 ± 0.2 | 0.28 ± 0.01 | 14.1 ± 0.4 | 0.38 ± 0.02 | 14.5 ± 0.2 | 0.34 ± 0.03 | 19.2 ± 0.1 | 0.28 ± 0.01 |
| Сентябрь | 8.9 ± 0.2 | 0.27 ± 0.02 | 9.7 ± 0.1 | 0.38 ± 0.03 | 9.4 ± 0.1 | 0.34 ± 0.02 | 9.2 ± 0.1 | 0.33 ± 0.02 |
| 2016 г. | ||||||||
| Июнь | 16.1 ± 0.7 | 0.23 ± 0.01 | 11.8 ± 0.8 | 0.22 ± 0.03 | 14.6 ± 0.8 | 0.26 ± 0.02 | Нет данных | Нет данных |
| Июль | 17.7 ± 0.4 | 0.24 ± 0.01 | 14.3 ± 0.3 | 0.23 ± 0.02 | 15.7 ± 0.4 | 0.26 ± 0.01 | ||
| Август | 14.9 ± 0.2 | 0.26 ± 0.02 | 13.6 ± 0.3 | 0.27 ± 0.03 | 13.7 ± 0.5 | 0.26 ± 0.02 | ||
| Сентябрь | 10.9 ± 0.1 | 0.24 ± 0.01 | 11.5 ± 0.2 | 0.31 ± 0.03 | 11.4 ± 0.2 | 0.24 ± 0.02 | ||
| 2017 г. | ||||||||
| Июль | 15.6 ± 0.4 | 0.27 ± 0.01 | 13.2 ± 0.5 | 0.39 ± 0.04 | 13.9 ± 0.5 | 0.34 ± 0.02 | Нет данных | Нет данных |
| Август | 14.9 ± 0.3 | 0.29 ± 0.02 | 13.6 ± 0.2 | 0.38 ± 0.03 | 13.8 ± 0.2 | 0.35 ± 0.01 | ||
| Сентябрь | 8.7 ± 0.2 | 0.35 ± 0.02 | 9.8 ± 0.1 | 0.56 ± 0.05 | 9.2 ± 0.1 | 0.47 ± 0.02 | ||
Максимальных значений влажность почвы достигает в ранневесенний период (первая декада мая) во время активного снеготаяния и в сентябре. По влажности почв ЭУ располагаются следующим образом: на нарушенном участке влажность в среднем составляет 0.31 ± 0.09 м3 м–3 (табл. 2); смешанный лес и сосняк зеленомошный характеризуются сходными величинами влажности почв – от 0.24 до 0.28 м3 м–3, а сосняк лишайниковый является наименее увлажненным из рассмотренных лесопокрытых ЭУ – влажность почвы в среднем составляет 0.21 ± 0.09 м3 м–3. Пик сезонной влажности для периода (июнь–сентябрь) всегда приходился на конец сезона – сентябрь. Для всех лесопокрытых ЭУ по условиям увлажнения почв существенно выделяется сезон 2012 г. – влажность почвы в июне–июле была в среднем на 60% ниже, чем в августе–сентябре.
Сезонная динамика почвенной эмиссии СО2
Сезонный ход почвенной эмиссии в исследованных типах земной поверхности имеет, как правило, типичный для лесных экосистем бореальной и умеренной зон вид – низкие скорости наблюдаются весной и поздней осенью, максимальные потоки приходятся на конец июля–начало августа (рис. 3), что обусловлено ходом температуры почвы и ее влиянием на активность автотрофов и гетеротрофов [16, 20, 27].
Рис. 3.
Сезонная динамика почвенной эмиссии СО2 для участков с разными типами растительного покрова за 5 лет измерений. Приведены средние значения со стандартными ошибками (N = 5).
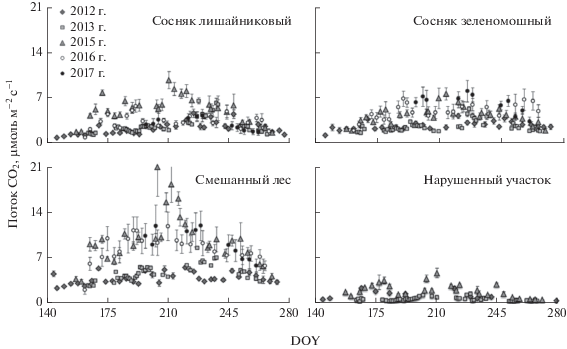
Средние величины почвенной эмиссии рассматриваемых ЭУ различались значимо (P < 0.01). Наименьшим эмиссионным потоком характеризуется нарушенный участок (1.1 ± 0.1 μмоль СО2 м–2 с–1), что обусловлено отсутствием растительного покрова и органических горизонтов почвы, основных источников продукции СО2. Известно, что ОВ почв на большой глубине характеризуется довольно слабой доступностью и не может обеспечить высокую скорость почвенной эмиссии, помимо этого с глубиной также снижается и активность гетеротрофов. При этом следует отметить, что при продолжительных засухах (например, 2012 и 2013 гг.) четко выраженной сезонной динамики эмиссий СО2 на ЭУ с нарушенной поверхностью не наблюдается, а средние потоки составляют лишь 0.40 ± 0.04 μмоль СО2 м–2 с–1 (2013 г.). Более выраженная сезонная динамика почвенной эмиссии отмечается во влажные годы (например, сезон 2015 г.), причем повышенные значения почвенного дыхания (до 4.4 μмоль СО2 м–2 с–1) наблюдались как во второй половине июня, так и в конце июля.
В сосняке лишайниковом средние потоки СО2 за вегетационный сезон составляли 3.4 μмоль СО2 м–2 с–1, варьируя от 0.9 до 9.8 μмоль СО2 м–2 с–1. Отметим, что для сосновых лесов южной тайги Центральной Сибири в искусственных насаждениях сосны ранее отмечены более низкие скорости почвенной эмиссии СО2 в течение вегетационного сезона – от 0.5 до 4 μмоль м–2 с–1 [28]. Причем авторы этой работы делают заключение о доминировании гетеротрофной составляющей в эмиссионном потоке СО2 из почвы.
В сосняке зеленомошном значения усредненных за сезон потоков СО2 близки к сосняку лишайниковому, составляя 4.3 μмоль СО2 м–2 с–1, варьируя от 1.4 до 8.2 μмоль СО2 м–2 с–1 в начале августа. В сосновых борах США [19, 26] рост почвенной эмиссии (до 12 μмоль СО2 м–2 с–1) также приходился на август и был связан с увеличением количества атмосферных осадков.
Максимальные значения почвенной эмиссии и ее вариации отмечены на участке со смешанным лесом (см. рис. 3): средние потоки СО2 за сезон значимо (P < 0.01) отличались от других типов участков и составляли 7.0 ± 1.2 μмоль СО2 м–2 с–1, варьируя от 1.8 до 20.9 μмоль СО2 м–2 с–1. Высокие потоки в данной экосистеме предположительно обусловлены не только гидротермическими условиями, но и ролью лесообразующих пород, разнообразием живого напочвенного покрова, а также активностью почвенных микроорганизмов. При сравнительной оценке почвенной эмиссии в темнохвойных и смешанных лесах Канады (Квебек) было установлено [29], что в смешанном лесу в течение вегетационного сезона почвенная эмиссия на 45% выше. Для смешанных лесов Франции [30] максимальные потоки достигают 10 μмоль СО2 м–2 с–1, однако пик потоков приходится на начало летнего сезона (июнь).
Межгодовая изменчивость эмиссии СО2
Рассмотрение межгодовой изменчивости позволяет судить о влиянии климатических факторов на изменения эмиссионного потока СО2 из почв при условии значительной вариации последних. В нашем случае изученные сезоны существенно различались по степени увлажнения, что и стало основным фактором, определяющим погодичную вариацию суммарной величины эмиссии СО2 из почвы.
Таблица 3.
Корреляция средних потоков за сезон (кг С м–2) каждого ЭУ с климатическими переменными. Полужирным выделены статистически значимые коэффициенты корреляции (p < 0.05)
| ЭУ | Коэффициент корреляции (в скобках значение p) | |||
|---|---|---|---|---|
| Tвозд, °С | Tпочв, °С | P *, мм | SWC, м3 м–3 | |
| Сосняк лишайниковый | –0.579 | 0.856 | 0.991 | 0.579 |
| (0.607) | (0.345) | (0.084) | (0.607) | |
| Сосняк зеленомошный | –0.127 | 0.999 | 0.813 | 0.127 |
| (0.919) | (0.033) | (0.396) | (0.919) | |
| Смешанный лес | –0.606 | 0.837 | 0.995 | 0.606 |
| (0.585) | (0.367) | (0.063) | (0.585) | |
| Нарушенный участок | –0.138 | 0.998 | 0.819 | 0.138 |
| (0.912) | (0.040) | (0.389) | (0.912) | |
Влияние климатических факторов на разных типах ЭУ отличается (табл. 3). Например, для сосняка лишайникового основное влияние на скорость почвенной эмиссии оказывает суммарное количество осадков за сезон, однако статистически эта связь незначима. Для сосняка зеленомошного отмечается наиболее высокая корреляционная связь с температурой почв (r = 0.999). В смешанном лесу на величину потока также самое существенное воздействие оказывает количество осадков. На участке с нарушенным напочвенным покровом основным фактором, определяющим эмиссию СО2, выступает температура почвы (r = 0.998). Заметим, что на всех ЭУ выявлена отрицательная зависимость от температуры воздуха, но статистически она незначима. Влажность почвы наиболее сильно отражается на потоках в сосняке лишайниковом и смешанном лесу – коэффициент корреляции составляет 0.579 и 0.606 соответственно.
Влияние типа экосистем было незначимым при P = 0.810 (табл. 4), но ЭУ значимо различались между собой в отдельные годы, т.е. главные факторы “тип экосистемы” и “год” оказались связанными (P < 0.010).
Таблица 4.
Результаты двух дисперсионных анализов: трехфакторного для скорости эмиссии СО2 и двухфакторного для годового потока СО2
| Факторы и их взаимодействия | Скорость потока СО2 | Годовой поток СО2 | ||||
|---|---|---|---|---|---|---|
| cтепени свободы | F | P | cтепени свободы | F | P | |
| Экосистема | 2 | 2.75 | <0.010 | 2 | 3.60 | 0.810 |
| Год | 4 | 6.52 | <0.001 | 4 | 10.12 | <0.001 |
| День | 10 | 46.35 | <0.001 | – | – | – |
| Экосистема × год | 8 | 2.18 | 0.010 | 8 | 2.12 | 0.008 |
| Экосистема × день | 20 | 2.86 | 0.032 | – | – | – |
| День × год | 40 | 3.11 | 0.045 | – | – | – |
| Экосистема × год × день | 80 | 2.55 | 0.002 | – | – | – |
Совокупность температур и условий увлажнения сезона статистически значимо (P < 0.05) влияет на величину потока (см. рис. 3). Исключение составляет нарушенный участок, где влияние было незначимым (P = 0.235).
В сосняке лишайниковом максимальные потоки отмечены в вегетационный сезон 2015 г. (см. рис. 3): средняя величина почвенной эмиссии составила 5.3 μмоль СО2 м–2 с–1, достигая 9.8 μмоль СО2 м–2 с–1 в середине сезона. В сосняке зеленомошном все сезоны по скорости почвенной эмиссии можно разбить на две группы по величине потоков СО2: 1-я группа – до 5 µмоль СО2 м–2 с–1; 2-я группа – от 5 до 10 µмоль СО2 м–2 с–1. К первой группе относятся два сезона – 2012 и 2013 гг., характеризующиеся дефицитом осадков, когда средняя за сезон скорость почвенной эмиссии составила 2.9 и 2.7 µмоль СО2 м–2 с–1 соответственно. Ко второй группе относятся сразу три сезона – 2015, 2016 гг. и вторая половина 2017 г., сезонные потоки СО2 в которой на 35–55% выше, чем в первой.
В засушливый сезон в сосновых лесах Бельгии [31] с моховым напочвенным покровом на песчаных почвах почвенная эмиссия достигала 2.3 μмоль м–2 с–1 в конце июля, при этом пик потоков совпадал с периодом увеличения количества атмосферных осадков.
ЭУ со смешанным лесом также, как и другие лесопокрытые участки, демонстрирует максимальные величины потоков СО2 в сезон 2015 г. (см. рис. 3): средние потоки СО2 за сезон сильно отличались от других типов участков и составляли 9.6 ± 0.7 μмоль СО2 м–2 с–1, достигая 20.9 μмоль СО2 м–2 с–1 в середине сезона 2015 г. Минимальные потоки отмечены в сезоны 2012 и 2013 гг. – среднесезонные значения потоков СО2 составили 3.7 и 4.5 µмоль СО2 м–2 с–1 соответственно.
Таким образом, существенные различия по величине почвенной эмиссии между экосистемами наблюдались во влажные сезоны (рис. 4). В условиях дефицита осадков в 2012 и 2013 гг. произошло уменьшение потока СО2 в среднем на 43%, а различия в величине эмиссионных почвенных потоков между лесными биогеоценозами нивелировались.
Рис. 4.
Поток СО2 с поверхности почвы за сезон измерений в 2012–2017 гг.: 1 – сосняк лишайниковый, 2 – сосняк зеленомошный, 3 – смешанный лес, 4 – нарушенный участок. Данные представляют собой средний поток со всех участков за вегетационный сезон ± стандартное отклонение (SD).

Наиболее высокие значения сезонной эмиссии составили в среднем 0.7 ± 0.1 кг С м–2 в 2015 г. В самый засушливый 2012 г. поток СО2 уменьшился в отдельных экосистемах в 2–2.5 раза по сравнению с 2015 г. Исследованные нами экосистемы оказались значительно более чувствительны к засухе, чем, к примеру, луговые экосистемы [32], для которых установлено, что поток СО2 из почвы снижается на 8–20% в зависимости от интенсивности и длительности засухи.
В более ранней работе [33] для сосняка лишайникового в районе исследований приводится величина экосистемного дыхания за 1998–2000 гг. порядка 372 г С м–2 год–1 (1364 г СО2 м–2 год–1). По оценкам А.В. Тимохиной [3], на основе данных обсерватории “ZOTTO” эмиссионный поток из экосистем зоны охвата высотной мачты варьировал в 2009–2013 гг. от 331 (2010 г.) до 398 г С м–2 сезон–1 (2012 г.), что согласуется с результатами проведенной нами работы. Согласно исследованию О.Б. Шибистовой с соавт. [16], на долю собственно почвенного дыхания приходится около 60% годовой величины экосистемного дыхания (23.1–23.4 моль м–2 год–1 ~ 280 г С м–2 год–1).
Зависимость почвенной эмиссии СО2 от температуры почвы
В более ранних исследованиях по сосновым лесам отмечалось [6], что при исключении эффекта засух при рассмотрении связи почвенной эмиссии с температурой зависимость (R2) между ними увеличивается с 0.06 до 0.59, становясь при этом значимой. Нами для изучения связи между почвенной эмиссией и температурой почвы было проведено разделение всех имеющихся данных на две группы по влажности почв (на сухой и влажный периоды). На рис. 2 представлено распределение коэффициента корреляции (r) при пошаговом удалении величин эмиссии в дни с высокими значениями влажности почв – от большей влажности к меньшей, и до последних трех пар значений. Точка пересечения двух зависимостей указывает на влажность, начиная с которой зависимость между потоком СО2 и температурой почвы становится значимой. Пороговая влажность (м3 м–3), разделяющая статистически значимые r от незначимых в ряду увеличения влажности и количества измерений, соответствует 0.30 для сосняка лишайникового, 0.14 – для сосняка зеленомошного, 0.24 – для смешанного леса и 0.23 – для нарушенного участка. Скорости почвенной эмиссии в сосняке лишайниковом, сосняке зеленомошном и смешанном лесу в группе с влажностью выше пороговой на 8, 25 и 14% соответственно больше, чем в группе, лимитированной содержанием воды (рис. 5).
Рис. 5.
Зависимость почвенной эмиссии СО2 от температуры почвы для двух групп по влажности почв (SWC – содержание воды в почве) на участках с разными типами растительного покрова: а – сосняк лишайниковый, б – сосняк зеленомошный, в – смешанный лес, г – нарушенный участок.

На всех лесопокрытых ЭУ при экстремально низкой влажности почв наблюдалась следующая тенденция: связь между потоком СО2 и температурой почвы была очень высокой, причем эта связь была статистически значимой (см. рис. 2). Подобная зависимость предположительно является проявлением стресс-реакции на условия полной засухи [15, 22].
Температурная чувствительность (Q10)
Температурную чувствительность минерализации почвенного ОВ выражают через коэффициент (Q10) [17], отражающий, во сколько раз увеличивается скорость этого процесса при повышении температуры на 10°С [30]. Коэффициент был рассчитан отдельно для двух выделенных групп влажности почвы (рис. 6).
Рис. 6.
Коэффициент Q10, рассчитанный для участков с разными типами растительного покрова. Данные приведены для двух групп по влажности почв: 1 – выше пороговой, 2 – ниже пороговой.

Температурная чувствительность автотрофного дыхания выше, чем у микроорганизмов [18, 34, 35], что обусловило более высокие значения Q10 для лесопокрытых участков. Для группы с влажностью выше порогового значения наибольшей величиной Q10 характеризуется смешанный лес (Q10 = 4.3). Коэффициенты Q10 в сосняках лишайниковом и зеленомошном сходны по величине и составляют 3.6 и 3.4 соответственно. На нарушенном участке (без растительного покрова) величина Q10 варьировала в зависимости от года от 1.5 до 1.8. Полученные значения Q10 для данного ЭУ близки к литературным оценкам для гетеротрофов (Q10 = 2 [7]), что подтверждает именно гетеротрофное происхождение выделяемого СО2.
В засушливые периоды температурная чувствительность у всех лесопокрытых экосистем находится на одном существенно более низком уровне (Q10 = 1.9). Наибольшие различия по температурной чувствительности отмечаются в сосняке лишайниковом – в группе с засушливыми условиями Q10 снижается на 65%, в смешанном лесу – на 58%, а в сосняке зеленомошном – на 43%. Минимальные различия по Q10 в двух группах с разными условиями увлажнения зафиксированы на нарушенном участке: в группе с засушливыми условиями почвенная эмиссия была на 18% ниже, чем в группе с повышенной влажностью.
Температурная чувствительность почвенной эмиссии меняется при различных условиях увлажнения. Для группы условно высокой влажности Q10 на всех лесопокрытых ЭУ характеризуется величинами в среднем на 55% выше, чем при засушливых условиях. Скорость эмиссии СО2 для нарушенного участка была менее чувствительна к росту температуры и, кроме того, Q10 слабо различался между влажными и сухими периодами.
ЗАКЛЮЧЕНИЕ
Суммарные потоки СО2 с изученных нами экосистем составляли от 0.3 до 0.8 кг С м–2 за сезон. Максимальными потоками характеризовался смешанный лес, а минимальными – нарушенный участок, при этом величина потоков в смешанном лесу была в среднем в 5 раз выше. Максимальных величин эмиссии почвенного СО2 достигают в конце июля–середине августа, однако в засушливые сезоны рост потоков наблюдался только со второй половины августа.
Климатические условия, и прежде всего условия увлажнения, оказывают существенное влияние на сезонный поток СО2, т.е. рассматриваемые нами лесные биогеоценозы Центральной Сибири, сформированные на хорошо дренируемых песчаных почвах, характеризуются значительной чувствительностью к засухе. Влияние типа леса проявляется только во влажные годы. Дефицит осадков в сезоны 2012 и 2013 гг. (54 и 65% от нормы соответственно) стал причиной уменьшения потока СО2 в среднем на 46% относительно сезонов с достаточным количеством осадков. Отдельные экосистемы в самый засушливый 2012 г. характеризуются значениями эмиссионных потоков СО2 в 2–2.5 раза ниже по сравнению с влажным 2015 г.
Работа выполнена при поддержке РФФИ (гранты № 17-05-01257, 18-34-00736).
Список литературы
Buchmann N. Biotic and abiotic factors controlling soil respiration rates in Picea abies stands // Soil Biology & Biochemistry 2000. V. 32. P. 1625–1635.
IPCC. Special report: global warming of 1.5°C. IPCC, Switzerland, 2018. 548 p.
Матвиенко А.И. Влияние азота на минерализацию углерода в почвах под лиственницей сибирской и сосной обыкновенной: Канд. дис. Красноярск, 2017.
Menyailo O., Huwe B.C. N-mineralization and denitrification as function of temperature and water potential in organic and mineral horizons of forest soil // J. of Plant Nutrition and Soil Science 1999. V. 162. P. 527–531. https://doi.org/10.1002/(SICI)1522-2624(199910)162
Raich J.W., Schlesinger W.H. The global carbon-dioxide flux in soil respiration and its relationship to vegetation and climate // Tellus B. 1992. V. 44(2). P. 81–99. https://doi.org/10.1034/j.1600-0889.1992.t01-1-00001.x
Wiseman P.E., Seiler J.R. Soil CO2 efflux across four age classes of plantation loblolly pine (Pinus taeda L.) on the Virginia Piedmont // Forest Ecology and Management. 2004. V. 192. P. 297–311. https://doi.org/10.1007/s10533-004-0566-3
Czimczik C.I., Schmidt M.W.I., Schulze E.-D. Effects of increasing fire frequency on black carbon and organic matter in Podzols of Siberian Scots pine forests // European J. of Soil Science. 2005. V. 56. P. 417–428. https://doi.org/10.1111/j.1365-2389.2004.00665.x
Lindroth A., Grelle A., More’n A.-S. Long-term measurements of boreal forest carbon balance reveal large temperature sensitivity // Global Change Biol. 1998. V. 4. P. 443–450. https://doi.org/10.1046/j.1365-2486.1998.00165.x
Lloyd J., Taylor J.A. On the temperature dependence of soil respiration // Functional Ecology. 1994. V. 8. P. 315–323. https://doi.org/10.2307/2389824
Grace J., Rayment M. Respiration in the balance // Nature. 1999. V. 404. P. 819–820. https://doi.org/10.1038/35009170
Rodeghiero M., Cescatti A. Main determinants of forest soil respiration along an elevation/temperature gradient in the Italian Alps // Global Change Biology. 2005. V. 11. P. 1024–1041. https://doi.org/10.1111/j.1365-2486.2005.00963.x
Sa’nchez M.L., Ozores M.I., Lo’pez M.J. et al. Soil CO2 fluxes beneath barley on the central Spanish plateau // Agricultural and Forest Meteorology. 2003. V. 118. P. 85–95. https://doi.org/10.1016/S0045-6535(02)00071-1
Schlesinger W., Andrews J. Soil respiration and the global carbon cycle // Biogeochemistry. 2000. V. 48. P. 7–20.
Davidson E.A., Janssens I.A. Temperature sensitivity of soil carbon decomposition and feedbacks to climate change // Nature. 2006. V. 440. P. 65–173. https://doi.org/10.1038/nature04514
Lee X., Wu H.J., Sigler J. et al. Rapid and transient response of soil respiration to rain // Glob. Change Biol. 2004. V. 10(6). P. 1017–1026. https://doi.org/10.1111/j.1529-8817.2003.00787.x
Shibistova O., Lloyd J., Evgrafova S. et al. Seasonal and spatial variability in soil CO2 efflux rates for a central Siberian Pinus sylvestris forest // Tellus. 2002. V. 54B. P. 552–567. https://doi.org/10.3402/tellusb.v54i5.16687
Kirschbaum M.U.F. Will changes in soil organic carbon act as a positive or negative feedback on global warming // Biogeochemistry 2000. V. 48(1). P. 21–51. https://doi.org/10.1023/A:1006238902976
Boone R.D., Nadelhoffer K.J., Canary J.D., Kaye J.P. Roots exert a strong influence on the temperature sensitivity of soil respiration // NATURE. 1998. V. 396. P. 570–572.
Irvine J., Law B.E. Contrasting soil respiration in young and old-growth ponderosa pine forests // Glob. Change Biol. 2002. V. 8(12). P. 1183–1194. https://doi.org/10.1046/j.1365-2486.2002.00544.x
Yuste J.C., Janssens I.A., Carrara A. et al. Interactive effects of temperature and precipitation on soil respiration in a temperate maritime pine forest // Tree Physiology. 2003. V. 23. P. 1263–1270.
Lia J.-T., Wang J.-J., Zeng D.-H. et al. The influence of drought intensity on soil respiration during and after multiple drying-rewetting cycles // Soil Biology and Biochemistry. 2018. V. 127. P. 82–89.https://doi.org/10.1016/j.soilbio.2018.09.018
Silva B.O., Moitinho M.R., Araújo Santos G.A. et al. Soil CO2 emission and short-term soil pore class distribution after tillage operations // Soil & Tillage Research. 2019. V. 186. P. 224–232. https://doi.org/10.1016/j.still.2018.10.019
Лесные экосистемы Енисейского меридиана / Под ред. Плешикова Ф.И. Новосибирск: Наука, 2002. 356 с.
Полосухина Д.А., Прокушкин А.С. Сравнительная характеристика запасов и изотопного состава почвенного органического вещества лесных биогеоценозов зоны охвата высотной мачты ZOTTO // Ломоносовские чтения на Алтае: фундаментальные проблемы науки и образования: Избр. труды междун. конф. Москва, 2017. С. 55.
Махныкина А.В., Прокушкин А.С., Ваганов Е.А. и др. Динамика потоков СО2 с поверхности почвы в сосновых древостоях Средней Сибири // Журн. Сибирского федерального ун-та. Биология. 2016. № 3. С. 338–357 [Makhnykina A.V., Prokushkin A.S., Vaganov E.A. et al. // J. of Siberian Federal University. Biology. 2016. V. 3. P. 338–357.] https://doi.org/10.17516/1997-1389-2016-9-3-338-357
Tang J., Misson L., Gershenson A. et al. Continuous measurements of soil respiration with and without roots in a ponderosa pine plantation in the Sierra Nevada Mountains // Agricultural and Forest Meteorology. 2005. V. 132. P. 212–227. https://doi.org/10.1016/j.agrformet.2005.07.011
Laganière J., Paré D., Bergeron Y. et al. The effect of boreal forest composition on soil respiration is mediated through variations in soil temperature and C quality // Soil Biology & Biochemistry. 2012. V. 53. P. 18–27. https://doi.org/10.1016/j.soilbio.2012.04.024
Матвиенко А.И., Макаров М.И., Меняйло О.В. Биологические источники почвенного СО2 под лиственницей сибирской и сосной обыкновенной // Экология. 2014. № 3. С. 182–188. [Matvienko A.I., Makarov M.I., Menyaylo O.V. Rus. J. of Ecology. 2014. V. 45(3) P. 174–180.] https://doi.org/10.7868/S0367059715020079
Van’t Hoff J.H. Lectures on Theoretical and Physical Chemistry // Chemical Dynamics. Part I. 1898. P. 224–229.
Vincent G., Shahriari A. R., Lucot E. et al. Spatial and seasonal variations in soil respiration in a temperate deciduous forest with fluctuating water table // Soil Biology & Biochemistry. 2006. V. 38. P. 2527–2535. https://doi.org/10.1016/j.soilbio.2006.03.009
Harper C.W., Blair J.M., Fay P.A. et al. Increased rainfall variability and reduced rainfall amount decreases soil CO2 efflux in a grassland ecosystem // Global Change Biology. 2005. V. 11. P. 322–334. https://doi.org/10.1111/j.1365-2486.2005.00899.x
Чебакова Н.М., Шульце Е.-Д., Выгодская Н.Н. и др. Энерго- и массообмен и продуктивность основных экосистем Сибири (по результатам измерений методом турбулентных пульсаций). 2. Углеродный обмен и продуктивность // Изв. РАН. Серия биол. 2014. № 1. С. 65–75 [Chebakova N.M., Schulze E.-D., Vygodskaya N.N. et al. Biology Bulletin. 2014. V. 1. P. 65–75.] https://doi.org/10.7868/S0002332914010044
Тимохина А.В., Прокушкин А.С., Онучин А.А. и др. Динамика приземной концентрации СО2 в среднетаежной подзоне Приенисейской Сибири // Экология. 2015. № 2. С. 110–119 [Timokhina A.V., Prokushkin A.S., Onuchin A.A. et al. // Rus. J. of Eco-logy. 2015. V. 2. P. 110–119.] https://doi.org/10.7868/S0367059715020122
Matteucci M., Gruening C., Goded Ballari I. et al. Components, drivers and temporal dynamics of ecosystem respiration in a Mediterranean pine forest // Soil Biology & Biochemistry. 2015. V. 88. P. 224– 235. https://doi.org/10.1016/j.soilbio.2015.05.017
Ruehr N.K., Buchmann N. Soil respiration fluxes in a temperate mixed forest: seasonality and temperature sensitivities differ among microbial and root-rhizosphere respiration // Tree Physiology. 2009. V. 30. P. 165–176. https://doi.org/10.1093/treephys/tpp106
Дополнительные материалы отсутствуют.