

Акустический журнал, 2021, T. 67, № 6, стр. 668-683
Вклад разных распределений активности ансамбля периферических волокон в формирование громкости и распознавание интенсивности тестовых импульсов, предъявляемых в тишине до и после помеховых импульсов
Л. К. Римская-Корсакова *
АО Акустический институт имени академика Н.Н. Андреева
117036 Москва, ул. Шверника 4, Россия
* E-mail: lkrk@mail.ru
Поступила в редакцию 12.08.2020
После доработки 17.06.2021
Принята к публикации 03.08.2021
Аннотация
С целью поиска физиологических основ слуховых эффектов повышения громкости и ухудшения распознавания интенсивности импульсов, предъявляемых до или после помеховых импульсов, а также эффекта ухудшения распознавания интенсивности одиночных импульсов моделировалась реакция ансамбля моделей периферических волокон. Полученная в ответ на однократное предъявление импульсов реакция ансамбля представлялась в виде двух комплементарных распределений: распределения вероятности появления спайков во времени и распределения межспайковых интервалов. В ответ на импульсные звуки такие распределения имели узкие области возбуждения. Для одиночных импульсов суммы спайков и межспайковых интервалов в областях возбуждения могли быть основой для формирования независимых “базовых” и “комплементарных” компонентов громкости импульса. Полученные в ответ на пару импульсов суммы спайков в двух областях первого распределения могли быть основой “базовых” компонентов громкости импульсов пары. Сумма межспайковых интервалов в первой области возбуждения второго распределения объединяла интервалы, соразмерные с длительностями каждого из импульсов пары, поэтому могла быть основой для “комплементарного” компонента громкости пары. Сумма межспайковых интервалов во второй области возбуждения объединяла интервалы, соразмерные с задержкой второго импульса относительно первого, и могла отвечать за выраженность периодичности пары. Величина “комплементарного” компонента громкости пары зависела от интенсивности и длительности импульсов пары, но не от положений и задержек этих импульсов. Учет “базовых” и “комплементарных” компонентов громкости позволил объяснить известные эффекты повышение громкости и ухудшения распознавания интенсивности одиночных и замаскированных импульсов. Специально проведенные слуховые эксперименты показали, что распознавание интенсивности замаскированных импульсов, по-видимому, вследствие влияния “комплементарного” компонента громкости пары, зависело от отношения интенсивностей импульсов в паре, но не от положений и задержек этих импульсов относительно друг друга.
1. ВВЕДЕНИЕ
Слуховой порог распознавания интенсивности звуков – это отношение наименьшего воспринимаемого приращения интенсивности dI к интенсивности тестового звука Is. Отношение dI/Is называют фракцией Вебера, а зависимость dI/Is от интенсивности тестового звука – функцией Вебера.
Слуховое распознавание интенсивности импульсных звуков, предъявляемых в тишине и условиях временной маскировки, имеет ряд особенностей, причины которых до сих пор не ясны. Перечислим эти особенности.
1. Полученная при действии одиночных тональных импульсов функция Вебера обнаружила горб при средних интенсивностях импульсов. Горб указывал на ухудшение распознавания интенсивности [1–5].
2. Если тональный импульс следовал за интенсивной шумовой помехой с задержкой в 100 мс, то функция Вебера также имела горб при средних интенсивностях импульсов [6–8].
4. Горб функции Вебера был больше, а распознавание интенсивности хуже, если тональный импульс со средней интенсивностью следовал с задержкой в 100 мс за более коротким помеховым импульсом [9].
3. Ухудшение распознавания интенсивности тональных импульсов со средними интенсивностями наблюдали, когда они следовали за или опережали помеховые импульсы [10–12]. При задержке в 100 мс горб функции Вебера был больше, если тестовый импульс опережал помеховый импульс.
5. В среднем диапазоне интенсивностей тональных импульсов, в котором возникал горб функции Вебера, регистрировали другой эффект – повышение громкости импульсов [12, 13]. Такой эффект выявлен сравнительно давно [14–16]. Громкость второго импульса в паре могла превысить громкость одиночного импульса, если интенсивность второго импульса была меньше интенсивности первого импульса, и задержка – меньше 200–400 мс. Эффект указывал на объединение громкостей импульсов пары, но явно отличался от эффекта временной суммации.
6. Горб функции Вебера и повышение громкости замаскированных импульсов возникали в различных конфигурациях слуховых раздражителей. Горб имели функции Вебера с аргументом интенсивности импульса Is, т.е. при варьировании интенсивности тестового импульса Is и постоянной интенсивности импульсной помехи Im [12, 13]. Горб также имели функции Вебера с аргументом интенсивности импульсной помехи Im, т.е. при варьировании интенсивности помехи Im и постоянной интенсивности тестового импульса Is [17].
Для объяснения перечисленных выше особенностей предложено большое число гипотез, учитывающих периферический и центральный анализ звуков. Самая первая гипотеза [6] предполагала, что горб функции Вебера был следствием разрыва между диапазонами кодирования импульсов волокнами слухового нерва (ВСН) с разной спонтанной активностью, что позже не подтвердили физиологические данные.
Есть мнение [18, 19], что свойства нелинейной компрессии базилярной мембраны [20] проявляются во многих слуховых явления. Однако свойства компрессии не часто рассматривали в качестве причины появления горба функции Вебера. На компрессию начали указывать после того, как интерес к проблеме горба заметно снизился [21–23]. Хотя моделирование периферической обработки звуков [24] показало, что за счет компрессии различия интенсивностей звуков на высоких частотах могли уменьшаться.
Однако сходства функций Вебера, полученных при разных положениях тестовых и помеховых импульсов, как и совпадение диапазонов интенсивностей импульсов, в которых возникали эффекты повышения громкости замаскированных импульсов и горб функций Вебера, скорее, указывали, что причины таких явлений связаны со свойствами не периферического кодирования, а процессами слухового центрального анализа.
С учетом когнитивных процессов и свойств памяти была предложена гипотеза ссылочного кодирования [10, 11]. Однако более убедительной представляется гипотеза [8, 12, 17] о том, что распознавание интенсивности импульсов, предъявляемых до и после помех, находится под влиянием интеграционных процессов формирования громкости импульсов.
Оценивая роль периферического кодирования, Мур указал [25], что частота разрядов ВСН является достаточной информацией для оценки распознавания интенсивности звуков в центральной нервной системе (ЦНС), но не исключил того, что ЦНС способна извлечь дополнительную информацию из активности ансамбля ВСН. Кариани [26] предположил, что слуховой нейронный код способен объединять информацию, заключенную в разных распределениях суммарной реакции ВСН, таких как распределение вероятности появления спайков во времени, распределение межспайковых интервалов, а также автокорреляционное распределение интервалов. В работе [27] моделировали реакцию ансамбля моделей ВСН, полученную в ответ на пару высокочастотных импульсов, которую представляли в виде указанных выше комплементарных распределений. Свойства таких распределений воспроизводили постстимульная гистограмма (ПСТГ), гистограмма распределения межспайковых интервалов (ГРМИ) и автокорреляционная гистограмма (АГ). Целью моделирования был поиск объяснений свойств распознавания интенсивности тестовых импульсов, предъявляемых до и после помеховых импульсов. Было показано, что изменения амплитуд импульсов вызывали заметные изменения ПСТГ и ГРМИ, но сравнительно слабые изменения АГ. Это указывало, что распознавание интенсивности могло быть под влиянием свойств ПСТГ и ГРМИ.
В данной работе путем моделирования искали обоснование гипотезы о влиянии процесса формирования громкости на распознавание интенсивности импульсов, предъявляемых в тишине, до и после помеховых импульсов. Исходили из того, что преобразование рецепторных потенциалов внутренних волосковых клеток в реакцию ансамбля ВСН, вызванную высокочастотными импульсами, является частью слухового кодирования интенсивности. Суммарная реакция ансамбля ВСН могла быть представлена в ЦНС в виде разных распределений, сохраняющих частотно-временные и амплитудные свойства импульсов. Такие распределения не могут формироваться на периферии, но могут в параллельных каналах ЦНС, по-видимому, участвующих в формировании громкости импульсов.
В работе моделировали реакцию ансамбля волокон слухового нерва в ответ на однократное предъявление одиночных и пар импульсов. Эту реакцию представляли в виде двух комплементарных распределений: распределения вероятности появления спайков во времени и распределения межспайковых интервалов, свойства которых воспроизводили ПСТГ и ГРМИ. Импульсы на гистограммах оставляли след в виде локальных зон возбуждения. Оценивали свойства таких возбуждений. Принимали, что суммы спайков или межспайковых интервалов в локальных областях могли быть основами для формирования разных компонентов громкости. Для оценки влияния этих компонентов на громкость и распознавание интенсивности импульсов вычисляли веса и индексы чувствительности d' к приращениям амплитуд импульсов по правилам теории обнаружения сигналов [28]. Для подтверждения справедливости выявленных в модельных экспериментах особенностей распознавания интенсивности амплитуд импульсов проводили слуховые эксперименты. Полагали, что совпадение результатов модельных и слуховых экспериментов подтвердит участие разных распределений активности ВСН в формировании громкости и распознавания интенсивности импульсов, а полученные данные расширят знания о временном нейронном коде [26], используемом слуховой системой для обнаружения и распознавания импульсных звуков.
2. МОДЕЛЬНЫЙ ЭКСПЕРИМЕНТ
2.1. Метод исследования: модели волокна слухового нерва и вычисляемые характеристики
Для моделирования реакции ансамбля ВСН была использована феноменологическая модель волокна слухового нерва (ВСН-модель) [29–31]. ВСН-модель имела несколько последовательных этапов преобразования входного сигнала X(t): (1) формирование реакции базилярной мембраны; (2) формирование рецепторного потенциала внутренней волосковой клетки и синаптического потенциала нейрона спирального ганглия; (3) генерация последовательности потенциалов действия (спайков) нейрона спирального ганглия.
На первом этапе, соответствующем формированию импульсного отклика базилярной мембраны Y(t), производили свертку сигнала Х(t) с импульсной характеристикой фильтра базилярной мембраны h(t):
Нелинейное преобразование сигнала на стадии механорецепции может воспроизвести сигмоидная функция [32, 33]. При моделировании реакций ВСН-моделей с разной спонтанной активностью (СА) мы использовали такие функции [31]. Рецепторный потенциал модели ВВК, R(t), полагали равным Φs(Y(t)), где наклон функции Φs зависел от СА так же, как наклоны характеристик вход‑выход реальных волокон [34]. В данном исследовании изучали реакции ансамбля волокон с низкой СА (~5 спайков за секунду), поэтому потенциал R(t) вычисляли согласно выражению:
Синаптический шум S(t) формировали как случайный процесс со средним значением и среднеквадратичным отклонением, пропорциональным потенциалу R(t). Чтобы воспроизвести СА, среднеквадратичное отклонение увеличивали на величину Z. При отсутствии сигнала (Х(t) = 0) среднее значение синаптического шума было равно нулю, а среднеквадратичное отклонение – Z. Формирование синаптического потенциала G(t) завершали путем интегрирования шума S(t): $G\left( t \right) = \int {{\text{exp}}\left( { - \frac{t}{\tau }} \right)S\left( t \right)dt} $, где τ – постоянная времени, равная 0.2 мс.
Преобразование потенциала G(t) в последовательность потенциалов действия (спайков) ВСН-моделей включало в себя сравнение G(t) с зависящей от времени пороговой функцией H(t). Если G(t) превышал H(t), то ВСН-модель генерировала спайк. Каждый спайк вызывал временное увеличение порога. В течение абсолютного рефрактерного периода ВСН-модель была не возбудимой. В течение относительного рефрактерного периода пороговая функция H(t) уменьшалась до своего начального значения H0. Функция H(t) в фазе относительной рефрактерности имела быструю и медленную компоненты. Начальные значения таких компонентов после генерации спайка увеличивались на остаточные значения компонентов, имевшие место до генерации спайков. Пороговая функция H(t) воспроизводила рефрактерные свойства, а также свойства адаптации, присущие реальным ВСН. Изменение пороговой функции H(t) после генерации спайка можно представить как:
Параметры ВСН-модели выбраны так, чтобы воспроизвести реакции реальных волокон слухового нерва [31]. Была учтена известная связь между морфологическими и физиологическими свойствами волокон с разной СА [34–39]. ВСН-модели с высокой СА, как реальные волокна, имели низкий порог срабатывания, крутую динамическую характеристику (зависимость средней частоты разряда от интенсивности тона) и узкий динамический диапазон. Они плохо воспроизводили огибающую амплитудно-модулированного сигнала и проявляли слабые свойства адаптации [40]. Напротив, ВСН-модели с низкой СА, как и реальные волокна, имели высокие пороги срабатывания, пологие динамические характеристики и широкий динамический диапазон. Они хорошо воспроизводили огибающую амплитудно-модулированного сигнала и обладали заметными свойствами адаптации [40]. ВСН-модель с адаптивным порогом хорошо воспроизводила известное несоответствие между спонтанной активностью, крутизной их динамической характеристики и способностью реальных волокон воспроизводить амплитудные модуляции. Причиной такого несоответствия были различия свойств рефрактерности и адаптации волокон, отвечающих за подстройку порога реакции волокна под входной потенциал G(t). Реакции ВСН-моделей также хорошо воспроизводили первично-подобные реакции и реакции типа “chopper”, встречающиеся у реальных звездчатых клеток вентрального кохлеарного ядра [29, 30].
Нелинейную компрессию на базилярной мембране [20] мы не учитывали, но изучали свойства преобразования аналоговых рецепторов внутренних волосковых клеток в последовательности спайков и межспайковых интервалов ансамбля ВСН.
Характеристическую частоту ВСН-модели определяла частота настройки фильтра базилярной мембраны, она была равна 4 кГц. Шаг вычисления в ВСН-модели составлял 1 мкс. В ответ на однократное предъявление одиночных или пар импульсов получали реакцию ансамбля из 100 ВСН-моделей. Реакцию представляли в виде постстимульной гистограммы (ПСТГ) и гистограммы распределения межспайковых интервалов (ГРМИ). Временная константа была равна 0.1 мс для ПСТГ и 100 мс для ГРМИ. Ширина бина в гистограммах составляла 0.1 мс. Тестовый и помеховый импульсы были отрезками синусоид длительностью 5, 10 или 15 мс с частотой в 4 кГц. Задержки между началами тестового и помехового импульсов были равны 20, 50 или 90 мс.
Полученные в ответ на одиночные импульсы ПСТГ и ГРМИ имели узкие области возбуждения [27]. Вычисляли сумму спайков, возникшую за время действия импульса в области возбуждения ПСТГ, и сумму интервалов, возникшую в областях возбуждения ГРМИ. Суммы спайков и интервалов полагали основами для “базового” и “комплементарного” компонентов громкости одиночного импульса.
ПСТГ и ГРМИ, полученные в ответ на пару, включающую помеховый и тестовый импульс, имели пары узких областей возбуждения. Суммы спайков в двух областях ПСТГ могли быть в основе “базовых” компонентов громкости импульсов пары. Области возбуждения ГРМИ сохраняли обобщенную информацию об импульсах пары. Сумма межспайковых интервалов в первой области возбуждения ГРМИ объединяла интервалы, соразмерные с длительностями каждого из импульсов пары, поэтому могла быть основой для “комплементарного” компонента громкости пары. Сумма межспайковых интервалов во второй области возбуждения ГРМИ объединяла интервалы, соразмерные с задержкой второго импульса относительно первого. При этом средний межспайковый интервал второй области мог отвечать за ощущение “высоты периодичности” пары импульсов [26, 41], а сумма интервалов – за выраженность или “силу высоты периодичности” [42].
Чтобы оценить участие разных компонентов громкости и силы высоты периодичности в распознавание амплитуд тестовых импульсов, был использован подход, предложенный в работе [43]. Для компонентов вычисляли индексы чувствительности d ' к приращению амплитуд импульсов. Индекс d ' представлял собой статистику, используемую в теории обнаружения сигнала [28], равную разности средних значений двух распределений, отнесенной к дисперсии распределений. При том, что одно из распределений характеризовало смесь сигнала и шума, а другое – только шум.
В данной работе, чтобы оценить значения индексов d ', были вычислены реакции ансамбля ВСН-моделей на две пары импульсов, в которых помеховый импульс имел амплитуду M, а тестовые импульсы – амплитуды S0 и (S0+ dS). Пару с амплитудой тестового импульса S0 принимали за “шум”, а пару с амплитудой тестового импульса (S0+ dS) – за “смесь сигнала и шума”. Реакциям ансамбля ВСН-моделей были присущи свойства распределения Пуассона, поэтому индексы чувствительности d ' для разных компонентов громкости определяли согласно выражениям (44):
За счет спонтанной активности ВСН-моделей индексы d' заметно варьировали в разных предъявлениях пар импульсов. Поэтому индексы вычисляли 10 раз и получали оценки средних значений и 90% доверительных интервалов.
В модельных экспериментах для разных компонентов громкости пар импульсов вычисляли психометрические функции как зависимости d '(N_S), d'(Max1) и d '(Max2) от приращения амплитуды импульса dS (в задаче распознавания амплитуд одиночных импульсов вычисляли d '(N_S) и d '(Max1), принимая амплитуду помехи M равной 0). Исходили их того, что при значении d ' = 0 порог обнаружения приращения dS определен с вероятностью в 50%; при d ' = 1 – 76%; при d ' = 3 – 98.5% [44]. Это значит, что при большей крутизне психометрической функции больше чувствительность компонента громкости к приращению амплитуды dS (т.е. меньше приращение амплитуды dS на пороге обнаружения с заданной вероятностью), лучше различимость (распознавание) амплитуд импульсов на основании такого компонента громкости.
Возникновение спайка или интервала рассматривали как “событие”. Сравнение чувствительностей компонентов громкости к приращению dS проводили в терминах “событий”. Индексы d ' сопоставляли с весами компонентов, равными суммам “событий”. Веса характеризовали громкость компонентов, а индексы d' – различимость амплитуд импульсов.
2.2. Результаты модельных экспериментов
2.2.1. Реакция ансамбля ВСН-моделей на одиночный импульс. На рис. 1 представлены реакции ансамбля ВСН-моделей, полученные в ответ на одиночный импульс. Импульс представлял отрезок синусоиды с частотой 4 кГц и длительностью 10 мс. Гистограммы получали на основании реакции ансамбля ВСН-моделей на однократное предъявление одиночного импульса. Локальные области возбуждения на гистограммах ПСТГ и ГРМИ отмечены как N_S и Max1, соответственно. Для этих областей были получены зависимости сумм спайков ΣN_S и интервалов от амплитуд импульсов S. По зависимостям определяли пороги обнаружения импульсов PS и PI, равные наименьшим амплитудам S, при которых возникали спайки и межспайковые интервалы.
Рис. 1.
Профили одиночного импульса (I), постстимульной гистограммы (II) и гистограммы распределения межспайковых интервалов (III); зависимости сумм спайков ΣN_S и межспайковых интервалов ΣMax1 в локальных областях возбуждения от амплитуды импульса S (IV). Параметр: амплитуда S. Абсцисса: время в мс (I, II), межспайковый интервал (МСИ) в мс (III), амплитуда S (IV), Ордината: суммы спайков (II) и межспайковых интервалов (III) в бине; сумма спайков или межспайковых интервалов в локальной области возбуждения (IV).

Согласно расчетам, пороги PS и PI были равны 100 и 300, соответственно. Учитывая значения этих порогов, амплитуды импульсов S были разделены на три группы, т.е. на низкие, средние и высокие. В этих группах методы кодирования импульсов ансамблем ВСН-моделей были разными.
Так, если S ≤ PI, то амплитуды импульсов S были низкими. При таких амплитудах сумма возникших спайков ΣN_S была меньше числа ВСН-моделей в ансамбле (ΣN_S < 100), а сумма межспайковых интервалов ΣMax1 была практически равна нулю. Ансамбль ВСН-моделей кодировал импульс числом возбужденных волокон, а метод кодирования был назван стохастическим.
Если S > PI, то амплитуды импульсов S были средними. При таких амплитудах все ВСН-модели в ансамбле генерировали более одного спайка (ΣN_S > ~100), но число моделей, в которых возникали межспайковые интервалы, было меньше общего числа волокон в ансамбле (0 < ΣMax1 < 100). Ансамбль ВСН-моделей кодировал импульс числом возбужденных волокон и числом сгенерированных спайков. Метод кодирования был смешанным, т.е. стохастическим и детерминистическим.
Если S $ \gg $ PI, то амплитуды S были высокими. При таких амплитудах суммы спайков и межспайковых интервалов превосходили общее число волокон в ансамбле (ΣN_S $ \gg $ 100 и ΣMax1 > 100). Ансамбль ВСН-моделей кодировал импульс преимущественно числом сгенерированных спайков. Такой метод кодирования был назван детерминистическим.
Возникшие в локальных областях возбуждения суммарные числа спайков ΣN_S и межспайковых интервалов ΣMax1 отвечали за формирование независимых “базового” и “комплементарного” компонентов громкости импульсов. Для оценки участия таких компонентов в формировании громкости и распознавании амплитуд импульсов, были сопоставлены веса компонентов ΣN_S и ΣMax1 (рис. 1, IV) и индексы чувствительности d '(N_S) и d '(Max1) (рис. 2). С увеличением амплитуды импульса S0 веса компонентов громкости ΣN_S и ΣMax1 повышались (рис. 1, IV), а значения индексов d '(N_S) и d '(Max1) и их вариабельность снижались (рис. 2). Вес базового компонента громкости (ΣN_S) всегда превышал вес комплементарного (ΣMax1) (рис. 1). Наибольшую чувствительность к приращению dS компоненты имели вблизи порогов, равных амплитудам импульсов, при которых возникали спайки или межспайковые интервалы (рис. 2).
Рис. 2.
Зависимости индексов чувствительностей d'(N_S) и d'(Max1) от приращения амплитуды импульса dS. Показаны средние значения индексов и 90% доверительные интервалы. Параметр – амплитуда стандартного импульса S0. Горизонтальные линии проходят через точки d' = 1 и 3.

При средних амплитудах импульсов (S0= 400 и S0 > PI) можно наблюдать максимальное несоответствие весов и чувствительностей компонентов. “Базовый” компонент ΣN_S с большим весом был менее чувствителен к приращению dS, а “комплементарный” компонент ΣMax1 с меньшим весом, наоборот, был более чувствительным. В случае сравнения амплитуд одиночных среднеамплитудных импульсов из-за несоответствия весов и чувствительностей разных компонентов громкости можно ожидать появления неопределенности в оценке громкости и поэтому ухудшение распознавания амплитуд импульсов.
При низких амплитудах импульсов (S0 = 250 и S0 ≤ PI) комплементарный компонент громкости не возникал. При высоких амплитудах импульсов (S0 = 750 и S0$ \gg $ PI) относительные различия весов и чувствительностей двух компонентов громкости снижались, и оба компонента в равной мере участвовали в распознавании амплитуд импульсов.
Следует отметить, что рассмотренные особенности кодирования импульсов не присущи кодированию продолжительных тонов. Расчеты показали, что вызванные тоном пороги PS и PI были сходными, поэтому средняя частота разряда у ВСН-моделей, как и у реальных волокон, была обратно пропорциональна среднему межспайковому интервалу. Метод кодирования тонов любых амплитуд был детерминистический, и комплементарный компонент не влиял на громкость и распознавание интенсивности тонов, как тот же компонент мог влиять на громкость и распознавание интенсивности импульсов.
Итоги раздела. Моделирование выявило различия периферического кодирования коротких импульсов с разными амплитудами. Изменения амплитуд импульсов вызывали изменения методов кодирования импульсов в ансамбле волокон, а также соотношений весов и чувствительностей к приращению амплитуд импульсов разных компонентов (сумм спайков и межспайковых интервалов) громкости импульса. Увеличение амплитуд импульсов сопровождали повышения весов компонентов громкости, но снижение чувствительности к приращению амплитуд импульсов. Наибольшая чувствительность компонентов была вблизи порогов появления спайков или межспайковых интервалов. Для импульсов пороги появления спайков и межспайковых интервалов были разными. Поэтому при средних амплитудах импульсов у разных компонентов громкости импульсов веса не соответствовали их чувствительности к приращению амплитуд. Можно предположить, что такое несоответствие могло ухудшить обнаружение пороговых приращений амплитуд или распознавание амплитуд импульсов.
2.2.2. Реакция ансамбля ВСН-моделей на пару импульсов. Рассмотрим влияние помехового импульса на веса и чувствительности компонентов громкости тестового импульса со средними амплитудами, а также пары импульсов.
Тестовый импульс в паре имел амплитуду S, а помеховый – амплитуду M (рис. 3). Амплитуды S были средними, равными 350–550. Анализировали изменчивость профилей ПСТГ и ГРМИ и характеристик ΣN_S, ΣN_M, ΣMax1 и ΣMax2 при варьировании амплитуд M в диапазоне 100–800, при разных длительностях импульсов, положениях и задержках D.
Рис. 3.
Профили пар тестовых и помеховых импульсов (I), постстимульные гистограммы (II) и гистограммы распределений межспайковых интервалов (III); зависимости сумм спайков ΣN_S, ΣN_M (IV) и межспайковых интервалов ΣMax1, ΣMax2 (V), возникших в локальных областях возбуждения, от амплитуды помехового импульса M. Амплитуда S тестовых импульсов была средней, она менялась в диапазоне от 350 до 550. Частоты импульсов равны 4 кГц. Длительности и задержки D между импульсами: (а, б) – 10 и 50 мс, (в) – 5 и 70 мс, (г) – 15 и 70 мс. На (а, б) – реакции на пары с разными положениями тестовых и помеховых импульсов. На (в, г) – реакции на пары с разными длительностями импульсов. Параметры на I–III: амплитуды тестовых S и помеховых M импульсов, их длительности и задержки. Абсцисса: время в мс (I, II), межспайковый интервал (МСИ) в мс (III), амплитуда помехового импульса M (IV, V). Ордината: амплитуды тестового S и помехового M импульсов (I); суммы спайков (II) и межспайковых интервалов (III) в бине; суммы спайков на (IV) и межспайковых интервалов (V) в локальных областях возбуждения.
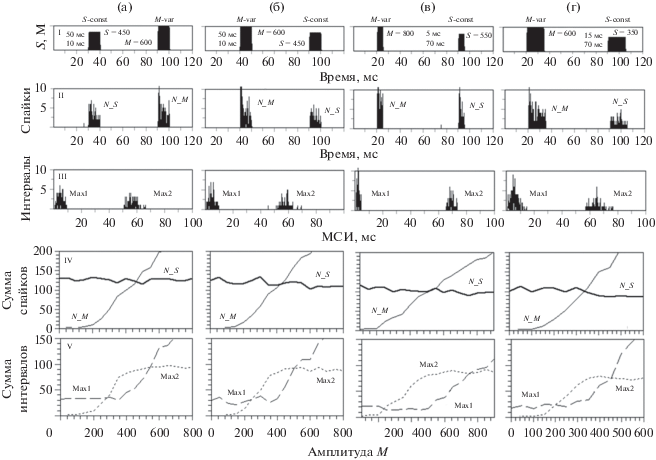
Положения локальных областей возбуждения на ПСТГ всегда соответствовали положениям тестовых импульсов и помех (рис. 3, II). Суммы спайков ΣN_S и ΣN_M были пропорциональны амплитудам S и M (рис. 3, IY). Если тестовый импульс следовал за помехой, то величина ΣN_S могла снизиться за счет свойств рефрактерности и адаптации ВСН-моделей.
Положения локальных областей возбуждения на ГРМИ не зависели от положений тестовых импульсов и помех (рис. 3, III). Область Max1 приходилась на интервалы, ширина которых не превышала длительности тестовых и помеховых импульсов, а область Max2 – на интервалы, ширина которых соответствовала задержке между импульсами пары. Изменения значений ΣMax1 и ΣMax2 при увеличении амплитуды M были сходными при разных положениях импульсов, разных задержках и длительностях импульсов в паре (рис. 3, V). Однако сами значения ΣMax1 и ΣMax2 зависели от отношения амплитуд S и M и длительностей импульсов. Сумма интервалов ΣMax1 была постоянной и низкой при M < S, но возрастала с увеличением амплитуды M при M ≥ S, а зависимость ΣMax1 от амплитуды M имела перегиб в точке M = S (рис. 3, V). Чем больше была длительность импульсов, тем быстрее росли ΣN_M и ΣMax1 с ростом М. Во всех случаях (рис. 3, V) сумма интервалов ΣMax2 была пропорциональна той амплитуде S или M, значение которой было меньше. Сумма ΣMax2 возрастала пропорционально росту амплитуды M при M < S, но была постоянной при M > S.
Мы принимали, что суммы спайков ΣN_S и ΣN_M могли быть в основе “базовых” компонентов громкости тестового и помехового импульсов, а суммы межспайковых интервалов ΣMax1 и ΣMax2 – “комплементарного” компонента громкости пары и компонента, отвечающего за “высоту периодичности” пары.
На рис. 4 сопоставлены веса компонентов ΣN_S, ΣN_M, ΣMax1 и ΣMax2 и индексы чувствительности d'(N_S), d'(Max1), d'(Max2) к приращению амплитуд тестовых импульсов dS. “Базовый” компонент громкости помехи был не чувствительным к dS. Тестовый импульс S0 имел среднюю амплитуду (400), а помеха M – низкую (250), среднюю (400) и высокую (750) амплитуды.
Рис. 4.
Схематичное представление тестового и помехового импульсов (I); зависимости весов разных компонентов громкости ΣN_S, ΣN_M, от амплитуды тестового импульса S = S0+ dS (II); зависимости индексов чувствительности d'(N_S), d'(Max1) и d'(Max2) от приращения амплитуды импульса dS (III). Частота и длительность импульсов равны 4 кГц и 10 мс, задержка между импульсами 50 мс. Помеховый импульс был (а) – последним или (б) – первым в паре. Остальное то же, что на рис. 2 и 3.

Изменения весов и чувствительностей разных компонентов были одинаковыми при разных положениях тестового и помехового импульсов в паре (рис. 4а и 4б). При большой скважности импульсов веса ΣN_S и индексы d '(N_S) “базового” компонента громкости тестового импульса не зависели от амплитуд помехи M и были сходными во всех представленных случаях. Однако вес ΣMax1 и индекс d '(Max1) “комплементарного” компонента громкости пары, а также вес ΣMax2 и индекс d '(Max2) компонента, отвечающего за периодичность пары, зависели от амплитуды помехи M (рис. 4). При увеличении M вес ΣMax1 повышался, а индекс d'(Max1) снижался. Индексы d '(Max2) были низкими при разных амплитудах M, и поскольку компонент Max2 отвечал за периодичность пары, мы полагали, что он не мог влиять на распознавание амплитуд тестовых импульсов.
Учитывая данные рис. 4, можно определить правила формирования громкости и распознавания интенсивности среднеамплитудных тестовых импульсов в присутствии помеховых импульсов c низкими, средними и высокими амплитудами (рис. 5).
Рис. 5.
Правила формирования громкости и распознавания интенсивности среднеамплитудного тестового импульса S, предъявляемого в присутствии помехового импульса M с низкими (S > M), средними (M ≥ S) и высокими (M $ \gg $ S) амплитудами. На I: схематичное представление зависимостей весов “базовых” компонентов громкости тестового ΣN_S и помехового ΣN_M импульсов, а также “комплементарного” компонента громкости пары импульсов пары ΣMax1 от амплитуды помехового импульса M. Абсцисса: амплитуды M, ордината: веса компонентов громкости. Овалы указывают на возможное объединение компонентов громкости. На II: для трех групп амплитуд помехи столбиками показаны громкости помехового М и тестового S импульсов. В столбиках отмечены вклады “базовых” компонентов громкости импульсов (белый цвет), и “комплементарного” компонента пары импульсов (серый цвет). Столбики темно-серого цвета соответствуют ожидаемым пороговым приращениям амплитуд тестовых импульсов (dS или dS') при распознавании амплитуд последних.

В формировании громкости помеховых и тестовых импульсов пары участвовали их “базовые” компоненты, но один “комплементарный” компонент громкости пары. “Комплементарный” компонент пары ΣMax1 мог дополнить тот “базовый” компонент ΣN_S или ΣN_M, вес которого были ближе к весу компонента ΣMax1. У пар с низкими (S > M) и высокими (M $ \gg $ S) амплитудами помеховых импульсов “комплементарный” компонент пары ΣMax1 дополнял более близкий по весу “базовый” компонент громкости помехового импульса ΣN_M (рис. 5). В таком случае за громкость тестового средне амплитудного замаскированного импульса отвечал только “базовый” компонент ΣN_S. Поэтому громкость такого замаскированного импульса могла быть меньше громкости одиночного импульса. Компонент ΣMax1 не влиял на обнаружение пороговых приращений dS (т.е. на распознавание амплитуд тестовых импульсов). Поэтому приращения dS могли быть близкими при низких и высоких амплитудах помеховых импульсов M.
Иная ситуация была у пар среднеамплитудных тестовых и помеховых импульсов (M ≥ S). Вес “комплементарного” компонента громкости пары ΣMax1 был ближе к весу “базового” компонента тестового импульса ΣN_S (рис. 5). При сходных индексах чувствительности компонентов (d '(Max1) ~ ~ d'(N_S)) (рис. 4) громкость среднеамплитудного замаскированного тестового импульса могла повыситься в результате сложения весов компонентов ΣN_S и ΣMax1. В формировании ΣMax1 участвовали оба импульса пары, поэтому громкость замаскированного импульса могла превысить громкость одиночного импульса. Если учесть, что при большем весе компонента меньше пороговое приращение амплитуд импульсов (рис. 2), то можно предположить, что пороговое приращение dS ' амплитуды замаскированных среднеамплитудных тестовых импульсов могло быть больше приращения dS замаскированных тестовых импульсов с низкими и высокими амплитудами (рис. 5, II). Повышение амплитуды среднеамплитудной помехи М могло привести: (1) к повышению веса компонента ΣMax1, (2) к повышению общей громкости тестового импульса, (3) к повышению порогового приращения dS ' амплитуды тестового импульса, т.е. ухудшению распознавания амплитуд импульсов.
Итоги раздела. В ходе моделирования было получено следующее. Две области возбуждения ПСТГ сохраняли информацию о величинах и положениях двух импульсов. Изменение амплитуды любого импульса вызывало изменение числа возникших спайков в соответствующей области возбуждения. Две области возбуждения ГРМИ несли в себе обобщенную информацию об импульсах пары. Изменения амплитуды любого импульса пары вызывали изменения сумм межспайковых интервалов в двух областях возбуждения. Положения областей возбуждения не зависели от положений, задержек и длительностей импульсов в паре. Учитывая свойства ПСТГ и ГРМИ, можно объяснить эффекты повышения громкости и ухудшения распознавания амплитуд среднеамплитудных тестовых импульсов, предъявляемых до и после импульсных помех (рис. 5). Согласно моделированию, ГРМИ не могла участвовать в формировании громкости и распознавании амплитуд импульсов, когда амплитуда тестового импульса S превышала амплитуду помехового импульса M (S > M) и когда амплитуда M значительно превосходила амплитуду S (M $ \gg $ S). Однако в диапазоне средних амплитуд тестовых и помеховых импульсов, когда амплитуда M была равна и несколько превышала амплитуду S (M ≥ S), ГРМИ могла влиять на громкость и распознавание. Можно предположить, что в этом случае “комплементарный” компонент громкости пары мог дополнить “базовый” компонент громкости тестового импульса, поэтому громкость тестового импульса могла повыситься. Повышение амплитуд помех M, соответствующих среднему диапазону, могло привести к пропорциональному повышению громкости, а также порогового приращения dS' амплитуды тестового импульса. Последний эффект не зависел ни от положений тестовых и помеховых импульсов, ни от задержек между ними (рис. 3).
3. ПСИХОАКУСТИЧЕСКИЙ ЭКСПЕРИМЕНТ
3.1. Цели и задачи слуховых экспериментов
Цель моделирования состояла в поиске особенностей периферического кодирования импульсных звуков, которые могли бы привести к инвариантности свойств распознавания амплитуд замаскированных тестовых импульсов к задержкам и временному положению помеховых импульсов, но зависимости распознавания от отношения амплитуд тестовых и помеховых импульсов.
Согласно данным моделирования (рис. 3, V), зависимость суммы межспайковых интервалов ΣMax1 от амплитуды помехового импульса M имела перегиб при равенстве амплитуд тестового S и помехового M импульсов (S = M). Мы полагали, что “комплементарный” компонент ΣMax1 громкости пары импульсов мог дополнять громкость тестового импульса и поэтому (рис. 2) ухудшать распознавание амплитуд тестового импульса в паре только выше точки перегиба, т.е. при M ≥ S (рис. 5).
Учет этих данных позволяет предсказать слуховую функцию Вебера как зависимость отношения минимального приращения интенсивности dI к интенсивности тестового импульса Is от интенсивности помехового импульса Im. У такой функции должны быть, по крайней мере, две характерные области, для случаев (1) Is > Im и (2) Im ≥ Is. При средних амплитудах тестовых и помеховых импульсов (при Im≥ Is) функция могла возрастать, обнаруживая ухудшение распознавания интенсивности, а на границе областей в точке, где Is = Im, профиль функции Вебера мог иметь перегиб. В области (1) ниже точки перегиба распознавание интенсивности тестового импульса могло быть наилучшим и не зависеть от Im. В области (2) выше точки перегиба распознавание могло ухудшаться пропорционально росту Im. При увеличении интенсивности тестового импульса (Is) точка перегиба функции Вебера с аргументом Im должна сместиться в сторону больших значений Im. Однако профили функции Вебера должны быть сходными при разных положениях тестовых и помеховых импульсов и разных задержках между ними.
Подтвердить справедливость предсказанного профиля слуховой функций Вебера могут результаты слуховых экспериментов. В массиве известных экспериментальных данных (см. Введение) есть указания на справедливость нашего предсказания. Так, Оберфельд [17] сравнивал громкость импульсов и распознавание интенсивности импульсов со средними амплитудами в 25, 55 и 85 дБ УЗД, которые следовали через 100 мс за помеховыми импульсами, интенсивности которых варьировали в диапазоне 0–90 дБ УЗД. Длительности и центральные частоты импульсов были равна 20 мс и 1 кГц. Если интенсивность помехи Im была ниже интенсивности стандартного импульса Is, то значения функции Вебера с аргументом Im флуктуировали относительно таковых, полученных в тишине. Однако в области Im≥ Is, когда интенсивность помехи Im превышала интенсивность стандартного импульса Is на 15–30 дБ, значения функции Вебера возрастали пропорционально интенсивности помехи Im, а громкость замаскированного импульса была больше громкости одиночного импульса.
С учетом данных работы [17] были проведены собственные слуховые эксперименты по распознаванию интенсивности импульсов, предъявляемых до и после импульсных помех. Цель измерений – показать зависимость профилей функций Вебера от отношения интенсивностей тестовых и помеховых импульсов пары, но независимость профилей от временного положения и задержек между импульсами пары.
3.2. Метод
Слуховые эксперименты проводили в заглушенной камере. Аппаратно-программный комплекс генерировал звуки, контролировал ход ведения эксперимента и регистрировал данные. Использовали адаптивную процедуру двух-альтернативного принудительного выбора. Испытательная последовательность имела два звуковых интервала продолжительностью 400 мс каждый. Пауза между интервалами была равна 500 мс. Один интервал содержал тестовый импульс и импульсную помеху, а другой – стандартный импульс и импульсную помеху. Звуки формировали в ходе эксперимента в виде файлов. Файлы воспроизводили в случайном порядке с использованием 24-битного цифроаналогового звукового адаптера EMU 0204 с частотой дискретизации 44.1 кГц. Звуки подавали на правое ухо слушателей через головные телефоны Sennheiser HD-265.
Для фиксации ответов слушателей использовали клавиатуру компьютера. Нажатием на клавиши клавиатуры слушатели сообщали, в каком из двух звуковых интервалов содержится тестовый импульс. В начале измерений интенсивность тестового импульса имела заведомо надпороговое значение. После двух правильных ответов испытуемых интенсивность тестового импульса уменьшали, а после одного неправильного ответа – увеличивали. Переход от уменьшения интенсивности к ее увеличению и наоборот называли точками реверсии.
Пороговое значение интенсивности тестового импульса определяли путем усреднения интенсивностей, полученных в последних восьми из одиннадцати точек реверсии. Полученный таким образом порог соответствовал порогу, определенному на основе психометрической кривой на уровне 71% [45]. Порог измеряли 4 раза. Если в этих измерениях ошибка среднего значения порога превышала 2 дБ, то проводили еще два дополнительных измерения. Пороговое значение интенсивности тестового импульса использовали для вычисления отношения Вебера, т.е. отношения dI/Is в дБ, где Is – интенсивность стандартного импульса; dI – пороговое приращение интенсивности стандартного импульса, равное разности интенсивностей стандартного и тестового импульсов.
Тестовые/стандартные импульсы, а также импульсные помехи имели синусоидальное заполнение с частотой 4 кГц и огибающую в виде гауссовой функции. Ширина полосы частот всех импульсов была равна 460 Гц (на уровне 0.7). Интенсивность стандартных и помеховых импульсов оценивали на основании их пиковых амплитуд, которые сравнивали с амплитудой непрерывного тона с частотой 4 кГц, определенной в дБ УЗД. Интенсивности стандартных и помеховых импульсов представляли в дБ УЗД и дБ над индивидуальным порогом слышимости (дБ ПС). Для этого для каждого слушателя измеряли пороги обнаружения одиночных импульсов и отношения Вебера, полученные при распознавании интенсивности одиночных импульсов. Последние отношения были самыми большими при интенсивностях одиночных стандартных тестовых импульсов Is в 20–30 дБ ПС.
В экспериментах были получены функции Вебера с аргументом Im, как зависимости отношения Вебера от интенсивности помехового импульса Im. Интенсивность Im варьировали в диапазоне 10–60 дБ ПС (~30–80 дБ УЗД). Параметрами были интенсивности стандартного импульса Is, равные 20, 30 или 40 дБ ПС (~40, 50 или 60 дБ УЗД), а также задержки D между серединами тестового/стандартного и помехового импульсов, равные 20, 50, 80 мс. Тестовый/стандартный импульс предъявляли как до, так и после помеховых импульсов.
В измерениях участвовали трое слушателей с нормальным слухом в возрасте до 35 лет. Среди них были две женщины (NUV, PNA) и один мужчина (MAA). Слушатели NUV и PNA уже участвовали в слуховых измерениях, слушатель MAA принимал участие в измерениях первый раз.
3.3. Результаты
Измеренные функции Вебера или зависимости порога распознавания интенсивности импульса (dI/Is) от интенсивности помехи (Im) представлены на рис. 6. Задержка D имела знак минус, если импульс опережал помеху, и плюс, если импульс следовал за помехой. Стандартные импульсы имели разные интенсивности (20, 30 и 40 дБ ПС). Задержка D была равна 50 мс.
Рис. 6.
Зависимости индивидуальных (MAA, NUV и PNA) и средних порогов распознавания интенсивности тестовых импульсов Is от интенсивности помеховых импульсов Im. Слева профили пар тестовых и помеховых импульсов. Помеховый импульс опережал или следовал за тестовым импульсом. Задержка D между импульсами равна 50 мс. Параметры: слушатель и интенсивность стандартного импульса в дБ ПС. Точки на оси ординат – индивидуальные и средние пороги распознавания интенсивности одиночных импульсов. Абсцисса: интенсивность Im в дБ ПС. Ордината: порог распознавания интенсивности dI/Is в дБ, где dI – пороговое приращение интенсивности тестового импульса.

Как показали измерения (рис. 6), индивидуальные и средние функции Вебера имели перегиб в точке, где интенсивность помехи была равна интенсивности стандартного импульса. При интенсивностях помех Im ниже точки перегиба, средние значения отношений Вебера соответствовали значениям, полученным в тишине, а при интенсивностях помех Im выше точки перегиба – возрастали пропорционально интенсивности помехи. При увеличении интенсивности стандартного импульса Is отношения Вебера, полученные в тишине и при наличии помех, снижались, а точка перегиба функции Вебера смещалась в сторону более высоких интенсивностей помех. Профили функций Вебера были сходными при разных положениях импульса относительно помехи.
На рис. 7 показаны функции Вебера, полученные для стандартных импульсов с интенсивностью в 40 дБ ПС и при задержках D в 20, 50 и 80 мс. Профили функций Вебера сохраняли рассмотренные на рис. 6 особенности при разных задержках D и разных положениях тестовых/стандартных импульсов относительно помеховых импульсов. Если интенсивность помехи была ниже, чем интенсивность стандартного импульса, то полученные средние значения отношений Вебера соответствовали значениям, полученным в тишине. Если интенсивность помехи была выше интенсивности тестового импульса, то отношения Вебера возрастали пропорционально интенсивности помехи. Диапазон интенсивностей помеховых импульсов, в котором функция Вебера возрастала пропорционально интенсивности помехи, составил ~20 дБ.
Рис. 7.
Зависимости индивидуальных (MAA, NUV и PNA) и средних порогов распознавания интенсивности тестовых импульсов Is от интенсивности помеховых импульсов Im. Интенсивность Is равна 40 дБ ПС. Задержки D между импульсами в паре равны ± 20, 50 и 80 мс. Остальное то же, что на рис. 6.

Итоги. Во-первых, полученные нами пороги распознавания интенсивности импульсов, предъявляемых до и после импульсных помех, соответствовали значениям, полученным в работе [17] (рис. 6, 7). Профили средних функций Вебера, имеющих интенсивность Im в качестве аргумента, зависели от отношений интенсивностей стандартных и помеховых импульсов. Профили имели точку перегиба, в которой интенсивность помехи Im была равна интенсивности стандартного импульса Is. Ниже точки перегиба отношения Вебера соответствовали значениям, полученным в тишине, а выше точки перегиба отношения Вебера возрастали пропорционально интенсивности помехи. При увеличении интенсивности стандартного импульса Is точка перегиба смещалась в сторону более высоких интенсивностей помех. Во-вторых, было показано, что профили функций Вебера не зависели ни от положений стандартных/тестовых импульсов относительно помеховых импульсов, ни от задержек между ними. В-третьих, свойства слухового распознавания интенсивности (рис. 6 и 7) соответствовали данным моделирования (рис. 3, V, рис. 4 и 5), учитывающим участие двух комплементарных распределений активности ансамбля периферических волокон в распознавании интенсивности.
4. ЗАКЛЮЧЕНИЕ
В данной работе методом моделирования искали объяснения следующих слуховых эффектов: (1) ухудшение распознавания интенсивности одиночных импульсов со средними интенсивностями; (2) повышение громкости и ухудшение распознавания интенсивности импульсов, предъявляемых до или после помех.
Для этого в модельном эксперименте анализировали методы кодирования высокочастотных звуков ансамблем периферических волокон, полагая, что процесс кодирования влиял на процесс формирования громкости звуков. Показано, что методы кодирования зависели от амплитуд и длительностей импульсов. Короткие импульсы могли быть закодированы ансамблем волокон тремя методами (стохастическим, смешанным и детерминистическим) в трех разных диапазонах амплитуд импульсов (низких, средних и высоких) (рис. 1). Стохастический метод кодирования низко амплитудных импульсов предполагал, что каждое из волокон в ансамбле реагировало на импульс только один раз, а реакция ансамбля волокон воспроизводила временной профиль импульса. Громкость импульсов могла формироваться с учетом числа сгенерированных спайков, равного числу возбужденных волокон в ансамбле.
Детерминистический метод высоко амплитудных импульсов предполагал, что каждое из волокон в ансамбле реагировало на импульс более одного раза. На генерацию спайков могли влиять рефрактерные свойства волокон, поэтому реакция ансамбля волокон не воспроизводила временной профиль импульса, но могла подчеркнуть периодичность (при наличии) огибающей. Громкость импульсов могла формироваться с учетом числа сгенерированных спайков в ансамбле, которое, практически, пропорционально числу возникших межспайковых интервалов.
В среднем диапазоне амплитуд за кодирование импульсов отвечал смешанный метод – стохастический и детерминистический. Чем короче был звук, тем шире был диапазон средних амплитуд. Громкость импульсов могла формироваться как с учетом числа возбужденных волокон, так и числа возникших межспайковых интервалов. Пороги появления межспайковых интервалов, всегда меньшие порогов появления спайков, определяли нижнюю границу средних амплитуд импульсов.
Изменения амплитуд импульсов сопровождали изменения методов их кодирования, способов формирования громкости и свойств распознавания амплитуд импульсов. Можно предположить, что в формировании громкости участвуют комплементарные распределения активности ансамбля ВСН (ПСТГ и ГРМИ). Суммарные числа спайков и межспайковых интервалов в узких областях возбуждения могли быть весами для “базового” и “комплементарного” компонентов громкости одиночного импульса. Участие таких компонентов в распознавании амплитуд импульсов оценивали по индексам d' чувствительности к приращениям амплитуд импульсов. Показано, что любые компоненты громкости с меньшим весом имеют наибольшую чувствительность вблизи порога появления спайка или межспайковых интервала (рис. 2). При средних амплитудах одиночных импульсов могло возникать несоответствие весов и чувствительностей к приращению амплитуд у разных компонентов громкости. Такое несоответствие могло быть причиной ухудшения слухового распознавания интенсивностей одиночных импульсов, которое наблюдали исследователи в многочисленных слуховых экспериментах [1–5].
Причины ухудшения распознавания амплитуд замаскированных импульсов были иными. Возникающие суммы спайков в двух узких областях возбуждения ПСТГ могли быть основами (весами) “базовых” компонент громкости импульсов пары. Сумма межспайковых интервалов ГРМИ, соответствующих длительности двух импульсов в паре, была комплементарным компонентом громкости пары. В диапазоне средних амплитуд тестовых и помеховых импульсов “комплементарный” компонент громкости пары мог дополнить “базовый” компонент громкости тестового импульса. Поэтому громкость тестового импульса повышалась, а его чувствительность к приращению амплитуды снижалась (рис. 2 и 5). Эффект мог иметь место при любых положениях тестовых и помеховых импульсов и разных задержках между ними (рис. 3 и 4).
Модельные данные позволили предсказать свойства слуховых функций Вебера, аргументом которых была интенсивность помехи Im. Свойства таких функций могли зависеть от отношений интенсивностей тестовых и помеховых импульсов, но не от положений тестовых и помеховых импульсов и задержке между ними.
Реальные слуховые функции Вебера с аргументами Is или Im действительно были сходными при разных положениях тестовых и помеховых импульсов и при задержках тестовых импульсов относительно помех в 100 мс [10–13, 17]. При этом было выявлено совпадение диапазонов интенсивностей импульсов Is или Im, в которых громкость импульсов пары превышала громкость одиночного импульса, а функция Вебера имела горб. Наши слуховые эксперименты подтвердили известные закономерности распознавания интенсивностей [17], а также обнаружили новые, предсказанные моделированием. Так, мы показали, что профили функций Вебера могли быть одинаковыми при разных задержках (20, 50 и 80 мс) тестовых импульсов относительно помеховых. За одинаковость мог отвечать “комплементарный” компонент громкости пары, временное положение которого не зависело от положений тестовых и помеховых импульсов. Вклад этого компонента в формирование громкости и распознавание интенсивности тестового импульса мог быть обнаружен, когда интенсивность помехи Im превышала интенсивность тестового импульса Is.
Таким образом, совпадение результатов модельных и слуховых экспериментов позволило подтвердить участие разных распределений активности ВСН в распознавания интенсивности импульсов, а полученные данные – расширить имеющиеся знания о временном нейронном коде [26], используемом слуховой системой для обнаружения и распознавания импульсных звуков.
Список литературы
Авакян Р.В., Радионова Е.А. Особенности дифференциальных порогов по интенсивности для короткого звукового сигнала // Акуст. журн. 1962. Т. 8. С. 407–411.
Raab D.H., Taub H.B. Click-intensity discrimination with and without a background masking noise // J. Acoust. Soc. Am. 1969. V. 46. № 4B. P. 965–968.
Carlyon R.P., Moore B.C. Intensity discrimination: a severe departure from Weber’s law // J. Acoust. Soc. Am. 1984. V. 76. 1369–1376.
Van Schijndel N.H., Houtgast T., Festen J.M. Intensity discrimination of Gaussian-windowed tones: indication for the shape of the auditory frequency-time window // J. Acoust. Soc. Am. 1999. V. 105. P. 3425–3435.
Римская-Корсакова Л.К. Слуховая периферическая адаптация в распознавании интенсивности коротких высокочастотных стимулов // Сенсорные системы. 2007. Т. 21. С. 294–306.
Zeng F.-G., Turner C.W., Relkin E.M. Recovery from prior stimulation. II. Effects upon intensity discrimination // Hearing Research. 1991. V. 55. P. 223–230.
Plack C.J., Viemeister N.F. The effects of notched noise on intensity discrimination under forward masking // J. Acoust. Soc. Am. 1992. V. 92. P. 902–1910.
Carlyon R.P., Beveridge H.A. Effects of forward masking on intensity discrimination, frequency discrimination, and the detection of tones in noise // J. Acoust. Soc. Am. 1993. V. 93. P. 2886–2895.
Schlauch R.S., Lanthier N., Neve J. Forward-masked intensity discrimination: Duration effects and spectral effects // J. Acoust. Soc. Am. 1997. V. 102. P. 461–467.
Plack C.J., Viemeister N.F. Intensity discrimination under backward masking // J. Acoust. Soc. Am. 1992. V. 92. P. 3097–3101.
Plack C.J., Carlyon R.P., Viemeister N.F. Intensity discrimination under forward and backward masking: Role of referential encoding // J. Acoust. Soc. Am. 1995. V. 97. P. 1141–1149.
Plack C.J. Loudness enhancement and intensity discrimination under forward and backward masking // J. Acoust. Soc. Am. 1996. V. 100. P. 1024–1030.
Zeng F.-G. Loudness growth in forward masking: Relation to intensity discrimination // J. Acoust. Soc. Am. 1994. V. 96. P. 2127–2132.
Irwin R.J., Zwisklocki J.J. Loudness effects in pairs of tone bursts // Percept. Psychophys. 1971. V. 10. P. 189–192.
Galambos R., Bauer J., Picton T., Squires K., Squires N. Loudness enhancement following con-tralateral stimulation // J. Acoust. Soc. Am. 1972. V. 52. P. 1127–1130.
Zwislocki J.J., Sokolich W.G. On loudness enhancement of a tone burst by a preceding tone burst by a preceding tone burst // Percept. Psychophys. 1974. V. 16. P. 87–90.
Oberfeld D. The mid-difference hump in forward-masked intensity discrimination // J. Acoust. Soc. Am. 2008. V. 123. P. 1574–1581.
von Klitzing R., Kohlrausch A. Effect of masker level on overshoot in running- and frozen-noise maskers // J. Acoust. Soc. Am. 1994. V. 95. P. 2192–2201.
Moore B.C., Oxenham A.J. Psychoacoustic consequences of compression in the peripheral auditory system // Psychol. Rev. 1998. V. 105. P. 108–124.
Robles L., Ruggero M.A. Mechanics of the mammalian cochlea // Physiol. Rev. 2001. V. 81. P. 1305–1352.
Nizami L. The intensity-difference limen for 6.5 kHz: an even more severe departure from Weber’s law // Percept. Psychophys. 2006. V. 68. 1107–1112. https://doi.org/10.3758/bf03193713
Pienkowski M., Hagerman B. Auditory intensity discrimination as a function of level-rove and tone duration in normal-hearing and impaired subjects: The “mid-level hump” revisited // Hearing Research. 2009. V. 253. P. 107–115.
Roverud E., Strickland T.A. Exploring the source of the mid-level hump for intensity discrimination in quiet and the effects of noise // J. Acoust. Soc. Am. 2015. V. 137. P. 1318–1335.
Heinz M.G., Colburn H.S., and Carney L.H. Rate and timing cues associated with the cochlear amplifier: Level discrimination based on monaural cross-frequency coincidence detection // J. Acoust. Soc. Am. 2001. V. 110. P. 2065–2084.
Moore B.C. An introduction to the psychology of hearing. 4th ed. Academic Press, London, 1997.
Cariani P.A. Temporal coding of periodicity pitch in auditory system: An Overview // Neural plasticity. 1999. V. 6. № 4. P. 147–171.
Римская-Корсакова Л.К. Проявления периферического кодирования в эффекте повышения громкости и особенностях распознавания интенсивности импульсов, предъявляемых до и после импульсных помех // Акуст. журн. 2017. Т. 63. С. 478–489.
Green D.M., Swets J.A. Signal Detection Theory and Psychophysics. New York, Wiley, 1966.
Дубровский Н.А., Римская-Корсакова Л.К. Определение параметров моделей слуховых нейронов, участвующих в выделении модуляций звуков: собственная периодичность модуляции // Акуст. журн. 1997. Т. 43. С. 421–428.
Дубровский Н.А., Римская-Корсакова Л.К. Определение параметров моделей слуховых нейронов, участвующих в выделении модуляций звуков: динамические и модуляционные передаточные характеристики // Акуст. журн. 1998. Т. 44. С. 173–179.
Римская-Корсакова Л.К., Дубровский Н.А., Телепнев В.Н. Проявление динамического кодирования амплитудно-модулированных звуков на уровне волокон слухового нерва // Росс. физиологический журн. им. И.М. Сеченова. 2003. Т. 6. С. 700–714.
Sachs M.B., Winslow R.L., Sokolowski B.H.A. A computational model of rate level functions from cat auditory-nerve fibres // Hearing Research. 1989. V. 41. P. 61–70.
Zagaeski M., Cody A.R., Russell I.J., Mountain D.C. Transfer characteristic of the inner hair cell synapse: Steady-state analysis // J. Acoust. Soc. Am. 1994. V. 95. P. 3430–3434.
Winter I.M., Robertson D., Yates G.K. Diversity of characteristic frequency rate-intensity functions in guinea pig auditory nerve fibres // Hearing Research. 1990. V. 45. P. 191–202.
Liberman M.C. Auditory nerve responses from cats raises in a low-noise chamber // J. Acoust. Soc. Am. 1978. V. 6. P. 442–455.
Kiang N.Y., Watanabe T., Thomas E.C., Clark L.F. Discharge patterns of single fibres in the cat’s auditory nerve. MIT Press, Cambridge, Massachusetts, 1965.
Joris P.X., Yin T.C.T. Responses to amplitude-modulated tones in the auditory nerve of the cat // J. Acoust. Soc. Am. 1992. V. 29. P. 215–232.
Sachs M.B., Abbas P.J. Rate versus level functions for auditory-nerve fibre in cats: tone burst stimuli // J. Acoust. Soc. Am. 1974. V. 56. P. 1835–1847.
Yates G.K., Manley G.A., Koppl C. Rate-intensity functions in the emu auditory nerve // J. Acoust. Soc. Am. 2000. V. 107. P. 2143–2154.
Frisina R.D. Subcortical neural coding mechanisms for auditory temporal processing // Hearing Research. 2001. V. 158. P. 1–27.
McClellan M.E., Small A.M. Pitch perception of pulse pairs with random repetition rate // J. Acoust. Soc. Am. 1967. V. 41. P. 690–699.
Plomp R. Auditory psychophysics // Annu. Rev. Psychol. 1975. V. 26. P. 207–232.
Viemeister N.F. Psychophysical aspects of auditory intensity coding. In: Auditory Function. Ed. Edelman G., Gall W., Cowan W. New York, Wiley, 1988. P. 213–241.
Egan J. Signal Detection Theory and ROC Analysis. New York, Academic Press, 1975.
Levitt H. Transformed up-down methods in psychoacoustics // J. Acoust. Soc. Am. 1971. V. 49. P. 467–477.
Дополнительные материалы отсутствуют.
Инструменты
Акустический журнал