

Агрохимия, 2022, № 7, стр. 55-65
Воздействие различных доз свинца на морфометрические, биохимические, цитогенетические показатели и продуктивность конских бобов (Vicia faba L.)
А. В. Дикарев 1, *, В. Г. Дикарев 1, Н. С. Дикарева 1
1 Всероссийский научно-исследовательский институт радиологии и агроэкологии
249032 Обнинск, Калужская обл., Киевское шоссе, 109 км, Россия
* E-mail: ar.djuna@yandex.ru
Поступила в редакцию 21.10.2021
После доработки 30.03.2022
Принята к публикации 15.04.2022
- EDN: VXARYW
- DOI: 10.31857/S0002188122070055
Аннотация
В вегетационном опыте с дерново-подзолистой супесчаной почвой были изучены морфометрические, цитогенетические, биохимические показатели и продуктивность конских бобов сорта Янтарные при действии свинца. Целью исследования была оценка воздействия токсического стресса и его отдаленных последствий, проявляющихся у сельскохозяйственных растений на примере конских бобов на основе вышеописанных показателей. Свинец вносили в виде Pb(NO3)2 (50, 100, 250, 500, 1000, 1500 мг/кг почвы) при закладке опыта, пробы листьев отбирали на 30-е сут и сохраняли в жидком азоте. Семена урожая, полученного с импактных растений в вегетационном опыте, использовали для анализа цитогенетических нарушений в апикальной меристеме корней проростков анафазным методом. Высота растений, измеренная на 30-е сут, изменялась не существенно до дозы 500 мг Pb2+/кг, далее наблюдали ее постепенное уменьшение. Свободный пролин в тканях растений накапливался в количествах, статистически значимо превышающих контроль, только при максимальной концентрации токсиканта. Содержание малонового диальдегида во всем исследованном диапазоне доз Pb2+ менялось в пределах контроля. Продуктивность растений (масса семян и их число) постепенно возрастала до 250 мг Pb2+/кг, затем (при дозах 500–1000 мг Pb2+/кг) величина этого показателя сокращалась и при 1500 мг/кг снижалась примерно до контрольного уровня. Митотический индекс (МИ) в изученном диапазоне доз Pb2+ изменялся несущественно, лишь при 1500 мг/кг наблюдался его рост. Кривая частоты профаз повторяла форму таковой для МИ, в то время как наблюдали накопление метафаз по мере возрастания концентрации Pb2+. Частоты ана- и телофаз оставались на близком к контролю уровне вплоть до дозы 500 мг Pb2+/кг, но уменьшались при дозах 1000 и 1500 мг Pb2+/кг. Частота аберрантных клеток (ЧАК) в апикальной меристеме клеток корней проростков демонстрировала нелинейный рост по мере возрастания дозы Pb2+. В диапазоне до 150 мг Pb2+/кг ЧАК возрастала несущественно, но в диапазоне 150–1000 мг/кг происходил значимый рост этого показателя, а при 1000–1500 мг/кг он выходил на плато. Основными типами нарушений были фрагменты, отставания хромосом и многополюсные митозы, а также иные аномалии, связанные с нарушением процесса цитокинеза. Был сделан вывод, что в рассматриваемом диапазоне доз свинца морфометрические и биохимические показатели растений изменялись мало, и значимый стресс проявлялся только при максимальных концентрациях. Тем не менее, токсический стресс оказывал существенное влияние на растения конских бобов и воздействовал на их следующее поколение, что проявлялось в увеличении цитогенетических нарушений в клетках проростков из семян импактных растений.
ВВЕДЕНИЕ
Свинец является одним из основных загрязнителей наземных и водных экосистем. Данный химический элемент, относящийся к группе тяжелых металлов (ТМ), поступает в окружающую среду в первую очередь в составе выхлопных газов, а также с отходами промышленности и сельского хозяйства [1]. В почвах сельскохозяйственных угодий содержание свинца может достигать 400–800 мг/кг, а в почвах промышленных зон – увеличиваться более 1000 мг/кг [2].
Накапливаясь в растениях в токсических концентрациях, свинец вызывает множественные повреждения во всех системах, органах, клетках, нарушает биохимические реакции, связанные с питанием, дыханием, фотосинтезом, изменяет активность ферментов и их состав, нарушает процессы минерального питания и водного обмена, меняет гормональный статус, воздействует на структуру плазмалеммы и ее проницаемость [2]. В результате на загрязненных этим ТМ почвах зачастую наблюдается снижение продуктивности сельскохозяйственных культур. Более того, продукты питания, произведенные на загрязненных территориях, могут содержать свинец в опасных для здоровья человека и домашних животных концентрациях [3].
Большинство исследований влияния ТМ на растения выполнено на модельных объектах. Это позволило оценить многие особенности реакции растений на воздействие наиболее опасных ТМ на клеточном, физиолого-биохимическом и молекулярно-генетическом уровнях. Несмотря на несомненную научную ценность и общебиологическую значимость полученной информации остается малоизученным большой пласт вопросов, касающихся особенностей действия конкретных ТМ на те или иные сельскохозяйственные культуры. Требуют дополнительного изучения механизмы поступления ТМ из почвы в растения, их транспорта, распространения и аккумуляции в разных органах и клеточных структурах, а также их влияние на продуктивность и качество урожая.
Учитывая насущную необходимость решения этих вопросов в условиях усиливающегося техногенного загрязнения сельскохозяйственных угодий, проводится широкая программа исследований различных аспектов действия основных ТМ на ряд сельскохозяйственных культур, затрагивающих разные уровни биологической организации растения. В настоящей работе в качестве объекта для такого исследования, посвященного оценке воздействия свинца на растительный организм, были взяты кормовые бобы (Vicia faba L.) – одна из важных кормовых культур. Цель работы – изучение цитогенетических и физиолого-биохимических механизмов воздействия свинца на вегетирующие растения конских бобов, оценка их продуктивности в условиях техногенного загрязнения, а также последствий токсического стресса для потомства этих растений.
МЕТОДИКА ИССЛЕДОВАНИЯ
Эксперимент по изучению влияния загрязнения почвы свинцом на морфометрические, цитогенетические, биохимические показатели и продуктивность конских бобов (Vicia faba L.) сорта Янтарные был заложен на дерново-подзолистой супесчаной почве. Ее агрохимические характеристики приведены в табл. 1.
Таблица 1.
Агрохимические характеристики дерново-подзолистой супесчаной почвы, использованной в опыте
| Показатель | Величина |
|---|---|
| pHKCl | 5.22 ± 0.01 |
| Гумус, % | 1.0 ± 0.01 |
| Hг , мг-экв/100 г почвы | 1.89 ± 0.02 |
| Сумма обменных оснований, мг-экв/100 г почвы | 5.3 ± 0.01 |
| Обменный K2O, мг/кг почвы (по Масловой) | 77.7 ± 1.3 |
| Подвижный P2O5, мг/кг почвы (по Кирсанову) | 127 ± 2 |
Растения выращивали в пластиковых сосудах по общепринятой методике [4]. Масса абсолютно сухой почвы в сосуде составляла 5 кг. При закладке опыта в почву вносили питательные вещества в виде водных растворов солей NH4NO3, KCl и KH2PO4 из расчета по N200P200K200 на 1 кг абсолютно-сухой почвы. Свинец добавляли в дерново-подзолистую почву в виде водного раствора соли Pb(NО3)2 в концентрациях 50, 100, 250, 500, 1000, 1500 мг/кг почвы. Поскольку свинец вносили в почву в виде нитрата, то количество добавляемого нитрата аммония корректировали в зависимости от дозы поллютанта так, чтобы в каждом случае общее количество азота было одинаковым во всех вариантах. Контролем служил вариант с NPK без внесения ТМ. Опыты проводили в четырехкратной повторности. Отбор проб растений для морфометрического и биохимического анализов проводили через 30 сут после появления всходов. Биохимические показатели – содержание малонового диальдегида (МДА) определяли по реакции с тиобарбитуровой кислотой [5], свободного пролина – по реакции с кислым нингидрином [6]. Для этих исследований на 30-е сут после появления всходов случайным образом отбирали листья, которые фиксировали в жидком азоте. После выполнения анализов для получения урожая оставляли по 8 растений/сосуд. После уборки урожая через 130 сут после всходов определяли структуру урожая по следующим показателям: высота растений, число бобов общее и число бобов с семенами, масса семян с сосуда, масса соломы, число семян с сосуда, масса 1000 семян.
Цитогенетический анализ клеток апикальной меристемы корней проростков бобов проводили после проращивания семян рулонным методом в сосудах с дистиллированной водой, помещенных в термостат при 24°С [7]. Для исследования брали семена бобов, полученные с растений, выращенных в ходе вегетационного опыта, чтобы оценить отдаленные эффекты воздействия свинца в изученном диапазоне концентраций. На 5–6-е сут проращивания в фиксаторе Кларка (этанол : уксусная кислота = 3 : 1) фиксировали корни длиной ≈10 мм. Для микроскопического исследования готовили временные давленые препараты. При этом корни мацерировали в растворе пектиназы (5 мкМ) в термостате при 37°С, а затем окрашивали ацетоорсеином (Merck, Германия) [8, 9]. Анализировали временные давленые препараты апикальных меристем на микроскопе Nikon Eclipse E200 (Япония). Рассчитывали объем выборки для цитогенетического анализа, обеспечивающий 95%-ный уровень статистической значимости; в среднем в каждом варианте анализировали 6–7 тыс. анафаз. При цитогенетическом анализе апикальных меристем определяли частоту аберрантных клеток, спектр и частоту цитогенетических аномалий (одинарных и двойных фрагментов и мостов, отставаний хромосом, мультиполярных митозов, нерасхождений хромосом, неправильных расхождений хромосом, патологических анафаз) [10–12]. Для оценки митотической активности клеток апикальной меристемы рассчитывали митотический индекс (МИ) для 12-ти препаратов, подсчитывали частоты клеток во всех 4-х фазах митоза; суммарно анализировали 1.0–1.5 тыс. клеток в каждом варианте.
Достоверность полученных результатов определяли путем расчета оптимального объема выборки, дисперсии, стандартных ошибок и отклонения, доверительного интервала и достоверности коэффициента линейной корреляции в программной среде MS Excel.
РЕЗУЛЬТАТЫ И ИХ ОБСУЖДЕНИЕ
Морфометрические параметры. Анализ морфометрических параметров и оценка внешнего вида растений кормовых бобов показали, что по мере увеличения внесенной в почву дозы свинца постепенно развивалось угнетение ростовых процессов, что выражалось в уменьшении высоты растений и задержке их развития. Однако даже при максимальной дозе поллютанта (1500 мг/кг) растения сохраняли возможность успешно завершить свой цикл вегетации, дав семена, и не демонстрировали визуально различимых признаков тяжелого стресса (появления морфозов, изменения окраски, исчезновения генеративных органов, завядания и др.). Таким образом, визуально наблюдаемые последствия токсического стресса были выражены в умеренной степени (рис. 1).
Рис. 1.
Внешний вид растений кормовых бобов сорта Янтарные (на сосудах указана внесенная в почву доза Pb2+, мг/кг).

Чтобы достоверно оценить изменение морфометрических параметров растений кормовых бобов по мере возрастания дозы свинца, рассматривали измененния такого показателя, как высота растений (рис. 2). Отмечено, что при относительно низких концентрациях ТМ в почве (50–250 мг/кг) происходили разнонаправленные изменения рассматриваемого показателя в пределах ошибки. В диапазоне от 250 до 500 мг/кг величина показателя стабилизировалась и практически не менялась, оставаясь близкой к контролю. При наибольших дозах Pb2+ высота растений постепенно уменьшалась, оказываясь при 1500 мг Pb2+/кг меньше контроля.
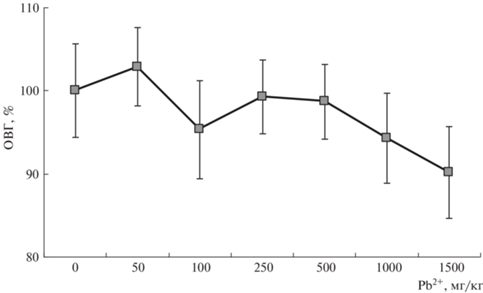
Биохимические параметры. Накопление МДА в листьях бобов демонстрировало тесную взаимосвязь с содержанием свободного пролина (рис. 3). Эта связь в данном случае имела существенное значение, т.к. первый из показателей характеризует уровень оксидативного стресса, а второй – реакцию защитной системы растения на стресс. В диапазоне доз ТМ до 250 мг Pb2+/кг происходила обычная в области малых доз любого стрессора [1, 13] разбалансировка стабильности окислительно-восстановительных процессов, выраженная в разнонаправленном изменении содержания МДА и пролина. В диапазоне доз 250–1000 мг Pb2+/кг в накоплении МДА и пролина наблюдали баланс, отражающий взаимодействие процессов повреждения и восстановления клеток растения. Повышение концентрации свинца до 1500 мг/кг нарушало сложившийся гомеостаз, отражением чего было увеличение содержания пролина. Это свидетельствовало об усилении процессов перекисного окисления липидов клеточных мембран в силу накопления свободных радикалов при данных дозах ТМ, и для нейтрализации избыточных количеств активных форм кислорода (АФК) растение увеличивало синтез защитного соединения – свободного пролина. Поэтому при максимальной дозе Pb2+ содержание пролина резко возрастало (до 1.6 раза от контроля).
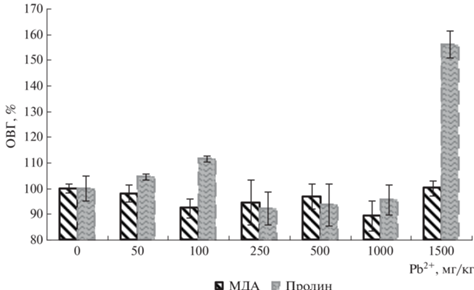
Рост содержания пролина при максимальной дозе ТМ позволил предположить, что концентрация 1500 мг Pb2+/кг была критической для кормовых бобов и при ней достигался предел устойчивости организма и происходило нарастание окислительного стресса. При дальнейшем возрастании дозы растения должны были демонстрировать серьезное угнетение в силу накопления АФК.
Структура урожая. Основные показатели продуктивности конских бобов представлены на рис. 4. Показано, что воздействие свинца в целом стимулировало продуктивность бобов, хотя этот эффект постепенно ослабевал при больших дозах ТМ, но даже при максимальной концентрации показатели не снижались ниже контроля. При этом урожай семян и их число демонстрировало выраженную реакцию на действие ТМ, а масса соломы и масса 1000 семян слабо реагировали на действие свинца.
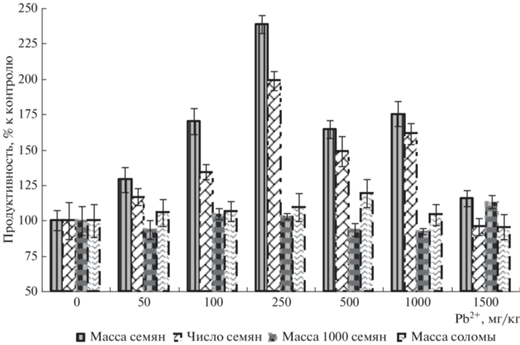
По главным показателям продуктивности – массе и количеству семян – получены вполне представительные дозовые зависимости, отражающие реакцию растений на действие свинца. В диапазоне низких доз ТМ (50–250 мг Pb2+/кг) отмечено возрастание массы семян почти в 2.5 раза по сравнению с контролем при концентрации 250 мг Pb2+/кг. Вероятно, этот феномен является следствием наличия сложной многоуровневой системы механизмов защиты растений. Однако, учитывая сложный продукционный процесс у этого семейства растений, когда формирование генеративных органов растянуто во времени и в пространстве (образование плодов происходит по мере роста стебля на разных уровнях, в разных условиях в отличии, например, от злаковых) наблюдаемое увеличение массы семян однозначно интерпретировать весьма сложно.
По мере возрастания концентрации свинца масса семян и их количество постепенно возрастали в диапазоне доз 50–250 мг Pb2+/кг почвы, но потом эти параметры постепенно снижались, не уменьшаясь, тем не менее, ниже контроля даже при максимальной дозе 1500 мг Pb2+/кг. Данную закономерность интересно сравнить с рис. 3, где отражены данные окислительно-восстановительного гомеостаза (ОВГ) растений. Содержание продукта перекисного окисления липидов (МДА) и защитного антиоксиданта (пролина) практически не менялось во всем диапазоне доз, но возрастало при максимальной дозе. Это и приводило к сокращению продукции семян, но, учитывая, что маркер стресса (МДА) оставался примерно на контрольном уровне, угнетения еще не развивалось. Потому масса семян не уменьшилась ниже контроля. Таким образом, наблюдали тесную взаимосвязь урожайности и активности окислительно-восстановительных процессов.
Масса соломы и масса 1000 семян практически не зависели от концентрации свинца. Интересно, что идентичность формы дозовых зависимостей этих показателей сильно отличалась от формы дозовой зависимости массы семян. Это свидетельствовало о том, что урожай семян в основном определяется их числом, что и подтверждается фактическими данными этого показателя и формой дозовой зависимости (рис. 4).
Цитогенетические параметры. Известно, что свинец, как и другие ТМ, обладает мутагенным потенциалом [14, 15]. Таким образом, токсический стресс, которому подверглось материнское растение, может сказаться на его семенном потомстве, вызвав повышенный выход нарушений генетического аппарата, и, как следствие, привести к угнетению растений 2-го поколения даже без реального наличия стрессора в среде. При этом быстрее всего таковые аномалии обнаружатся в клетках образовательных тканей растения, поскольку меристемы, активно развиваясь, состоят из множества молодых и делящихся клеток. А поскольку для нормального развития растения необходимо поглощение минеральных веществ из почвы в достаточном количестве, то состояние апикальной меристемы корня имеет критическое значение для дальнейшего состояния даже уже взрослого растительного организма. Это связано со значимостью транспортной и барьерной функций корня в поддержании гомеостаза растительного организма. Поэтому для оценки состояния образовательных тканей проростков бобов был использован цитогенетический анализ, позволяющий оценить частоты мутационных событий в клетках. Для этой работы были взяты семена бобов, выращенные в процессе вегетационного эксперимента на импактных и контрольных растениях.
В результате проведенного исследования показано, что митотический индекс (МИ) в диапазоне доз 50–1000 мг Pb2+/кг менялся в пределах ошибки (рис. 5а), однако при максимальной концентрации (1500 мг Pb2+/кг) отмечен его некоторый рост. При этом главным образом отмеченный рост МИ при максимальной дозе ТМ определялся возрастанием частоты профаз, и кривая МИ по форме была практически идентична таковой для профаз. Такой эффект можно объяснить тем, что по мере возрастания концентрации ТМ постепенно накапливались цитогенетические нарушения, которые приводили к торможению процесса пролиферации на стадии профазы. Сходные явления отмечены нами в предшествующих исследованиях [16–20].
Рис. 5.
Митотическая активность клеток апикальной меристемы корней проростков кормовых бобов: (а) – митотический индекс и частота профаз. (б) – частоты мета-, ана- и телофазы, ВК – общее количество клеток.
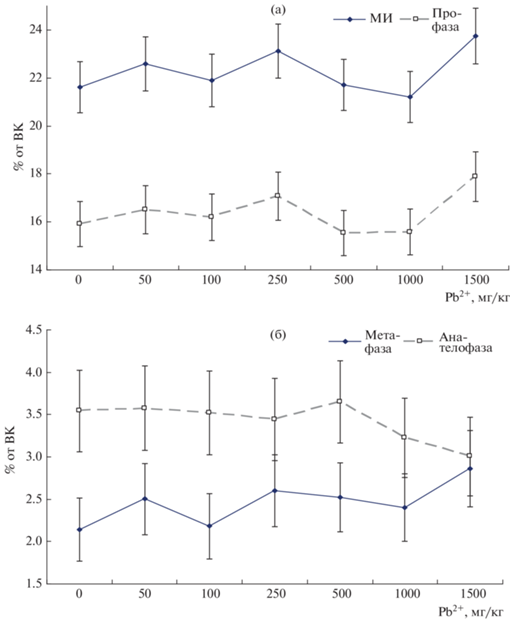
Впрочем, рассмотренные дозы оказались недостаточными для развития серьезного токсического стресса, который мог привести к полному угнетению процесса пролиферации. Поэтому, как это показано на рис. 1, даже при максимальной дозе Pb2+ растения бобов, хотя и демонстрировали заметное угнетение, но все же были еще способны более или менее успешно развиваться, входя в фазу цветения и давая семена. Эти выводы подтверждаются также и данными рис. 5б, где показано, что в целом частоты остальных фаз митоза не уменьшались значимо ниже контроля даже при максимальной дозе свинца (1500 мг/кг).
Однако по мере возрастания концентрации свинца происходило постепенное накопление метафаз (рис. 5б), при этом частоты ана- и телофаз, оставаясь стабильными до дозы 500 мг/кг, после этого (при 1000 и 1500 мг Pb2+/кг) начинали почти линейно уменьшаться. Указанные наблюдения можно объяснить тем, что, вероятно, на стадии метафазы при высоких концентрациях свинца развивалась еще одна блокировка митотического процесса. Это вело к накоплению метафаз при сокращении частот заключительных фаз митоза. Поскольку для успешного осуществления ана- и телофаз критически важна правильная работа аппарата веретена деления, можно предполагать, что свинец инициировал повреждение этой структуры.
Помимо оценки митотической активности, цитогенетический анализ клеток апикальных меристем корней проростков конских бобов позволил определить основные параметры концентрационной зависимости частоты аберрантных клеток (рис. 6). Концентрационная зависимость частоты аберрантных клеток имела характерную нелинейную форму, аналогичную той, что наблюдали в наших исследованиях с другими ТМ и для других растений [14–20]. Этот феномен был детально исследован и убедительно объяснен в работе [13]. Основной причиной нелинейности было действие ряда механизмов защиты растений от токсического воздействия, в данном случае, ТМ. Однако эта закономерность имеет значительно более широкое распространение, т.к. основные особенности этого явления были обнаружены при исследовании радиационного воздействия на растения [21]. В настоящем исследовании концепция нелинейного ответа организма на токсическое воздействие [13, 21, 22] нашла очередное подтверждение. Недостаточный набор концентраций фактора воздействия (Pb2+) не позволил, к сожалению, выявить подробности этой дозовой зависимости. Однако даже в этих условиях обнаружено резкое возрастание частоты аберрантных клеток уже при умеренных концентрациях токсиканта в диапазоне 100–500 мг Pb2+/кг. Дальнейшее повышение концентрации свинца почти не вызывало увеличения величины этого показателя, который практически выходил на плато в диапазоне доз 1000–1500 мг Pb2+/кг. Это могло свидетельствовать об общности процессов, происходящих в растительном организме при действии ксенобиотиков разного типа. Например, в апикальных меристемах конских бобов наиболее массовым типом цитогенетических нарушений (табл. 2) при действии свинца были отставания хромосом. Более того, частота этих нарушений статистически значимо коррелировала с концентрацией ТМ (коэффициент корреляции у бобов равен 0.965). Такого же уровня корреляционная связь отмечена и для интегрального параметра цитогенетических нарушений как частота аберрантных клеток (коэффициент корреляции равен 0.900).
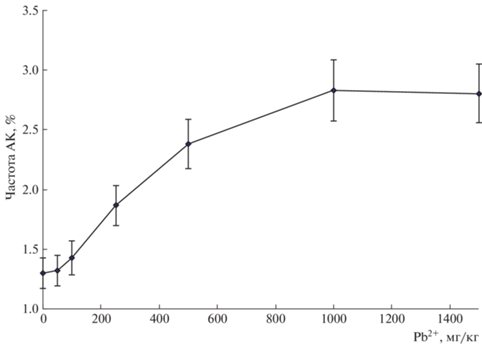
Таблица 2.
Спектр цитогенетических нарушений в апикальной меристеме корней проростков конских бобов
| Доза Pb2+, мг/кг почвы | ЧДК | ЧАК | Частота нарушений отдельных типов, % | |||||||
|---|---|---|---|---|---|---|---|---|---|---|
| f ' | f '' | m' | m" | g | mp | ndis | and | |||
| 0 | 7610 | 1.30 | 0.094 | 0.422 | 0.187 | 0.047 | 0.609 | 0 | 0 | 0 |
| 50 | 7640 | 1.32 | 0.049 | 0.247 | 0.198 | 0.148 | 0.642 | 0 | 0 | 0 |
| 100 | 6930 | 1.43 | 0.039 | 0.270 | 0.077 | 0.116 | 0.771 | 0.077 | 0 | 0.116 |
| 250 | 6530 | 1.87 | 0.043 | 0.522 | 0.087 | 0.043 | 1.087 | 0.174 | 0 | 0.087 |
| 500 | 5290 | 2.38 | 0.103 | 0.514 | 0.171 | 0.651 | 1.234 | 0.069 | 0.034 | 0.274 |
| 1000 | 4210 | 2.83 | 0.000 | 0.468 | 0.156 | 0.468 | 1.768 | 0.104 | 0.104 | 0.156 |
| 1500 | 4640 | 2.81 | 0.043 | 0.510 | 0.128 | 0.255 | 1.872 | 0.255 | 0.043 | 0.170 |
| r (доза) | 0.900* | –0.399 | 0.569 | –0.094 | 0.467 | 0.965* | 0.774* | 0.733* | 0.577 | |
Примечание. ЧДК – число делящихся клеток, ЧАК – частота аберрантных клеток, f ' – фрагмент одинарный, f " – фрагмент двойной, m' – мост одинарный, m" – мост двойной, g – отставание, mp – многополюсный митоз, pata – патологическая анафаза, and – неправильное расхождение хромосом, r – коэффициент линейной корреляции (знаком “*” помечены данные, достоверные при р ≤ 0.05).
Полученные в нашем исследовании зависимости частоты аберрантных клеток в апикальной меристеме бобов от содержания свинца в почве сходны по форме с полученными ранее дозовыми зависимостями на проростках ячменя при γ-облучении [13, 14, 21, 22] и действии свинца [16, 17]. Было показано, что между концентрацией свинца в почве и ЧАК, а также частотой суммы аберраций всех типов, частотой нарушений хромосом, частотой нарушений митотического аппарата существует статистически значимая корреляционная связь. Это свидетельствовало о том, что воздействие свинца затрагивает в первую очередь генетические структуры клеток растений.
Кроме традиционных нарушений, регистрируемых анафазным методом, таких как мосты, фрагменты, отставания хромосом и редко встречающиеся мультиполярные, чаще всего, трехполюсные митозы, свинец вызывал задержку митотического цикла на стадии профазы, нерасхождение и неправильное расхождение хромосом, образование патологических анафаз. Повышенный выход всех этих нарушений также отмечен нами.
Цитогенетический анализ клеток апикальных меристем корней проростков показал, что по мере роста дозы свинца происходило накопление цитогенетических аномалий, которые в конечном счете могут вести к подавлению процесса пролиферации. Изученный диапазон доз был недостаточен для того, чтобы остановить митоз, однако при максимальной дозе уже наблюдали тенденцию к этому. На это указывало накопление профаз в ущерб остальным фазам клеточного деления. Описанные эффекты аналогичны тем, о которых сообщали другие исследователи [14, 15] при изучении действия радиации на растения. Появление в клетках многополюсных митозов свидетельствовало о повреждении веретена деления, ведущего к остановке процесса пролиферации в метафазе. Таким образом, свинец повреждает митотический аппарат и оказывает мутагенное действие на клетки. В работе [10] описаны такие нарушения при действии колхицина и других ядов.
Достаточно высокая частота двойных мостов, низкая – одинарных мостов и невысокие частоты одинарных фрагментов могли свидетельствовать о том, что свинец воздействует на ДНК главным образом в фазе G2 клеточного цикла. Однако наблюдавшаяся нами и другими исследователями задержка деления в профазе показала, что свинец также нарушает процесс репликации ДНК [10–12, 18]. Эти эффекты нельзя считать общим механизмом токсического действия металлов, поскольку внесение в среду меди или алюминия в концентрации, подавляющей рост корней в сравнимой степени, не вызывало аналогичных изменений в структуре микротрубочек. Повреждение микротрубочек является характерной особенностью токсического действия свинца на растения [11, 12]. По нашим данным, при максимальной дозе свинца большинство всех нарушений было связано именно с повреждениями аппарата веретена деления и микротрубочек, что позволило рассматривать подобное в качестве маркера цитогенетического воздействия свинца.
ЗАКЛЮЧЕНИЕ
Таким образом, установлено, что в процессе вегетации морфометрические параметры растений кормовых бобов в изученных диапазонах концентраций свинца слабо реагировали на воздействие тяжелого металла (ТМ). Изменения внешнего вида и высоты растений были заметными только при высоких дозах. При действии свинца в диапазоне концентраций до 1000 мг/кг почвы содержание МДА и пролина в листьях конских бобов статистически значимо не отличалось от контроля; статистически значимое увеличение содержания пролина более чем в 1.6 раза по сравнению с контролем было отмечено лишь при концентрации 1500 мг Pb2+/кг почвы. Из основных показателей продуктивности наиболее выраженными были изменения массы и числа семян, которые во всем исследованном диапазоне доз свинца превышали контроль. Величины этих показателей постепенно возрастали до дозы 250 мг Pb2+/кг (в 2.0–2.5 раза по сравнению с контролем), а потом начинают уменьшаться, но и при 1500 мг Pb2+/кг не были меньше контроля. Масса 1000 семян и соломы в этом диапазоне доз статистически значимо не изменялась.
Митотический индекс при действии свинца во всем диапазоне доз оставался в основном стабильным, однако при максимальной дозе 1500 мг Pb2+/кг отмечен некоторый его рост за счет накопления профаз. При этом также постепенно возрастала и частота метафаз, особенно при дозах 1000 и 1500 мг Pb2+/кг, в то же время частоты ана- и телофаз при этих дозах свинца уменьшались. Цитогенетические нарушения в апикальной меристеме корней проростков конских бобов были обнаружены уже в диапазоне низких доз ТМ, являясь наиболее надежными и чувствительными показателями, позволившими оперативно обнаруживать и детально охарактеризовать реакцию растений на действие свинца.
Основным выводом по результатам исследования можно считать следующее. В рассматриваемом диапазоне доз свинец еще не вызывает у растений конских бобов развития значимого токсического стресса. Это выражается в том, что морфометрические, биохимические параметры и продуктивность лишь при максимальной дозе Pb2+ демонстрируют некоторые отличия от контроля. Однако перенесенное воздействие не проходит бесследно, оказывая мутагенный эффект на растения, который обнаруживается и во втором поколении через повышенный выход цитогенетических аномалий в образовательных тканях проростков.
Таким образом, анализ частоты и спектра цитогенетических нарушений позволил оценить механизмы действия определенных токсикантов на растения разных видов прежде, чем стрессовые эффекты приобретают визуально заметное проявление или сказываются на изменении биохимических параметров. Следовательно, цитогенетические методы могут быть использованы для идентификации действия конкретных ксенобиотиков и помочь в установлении общих и индивидуальных особенностей этих воздействий.
Собранные в результате проведенного исследования данные могут быть полезны для выработки методики анализа эффектов, оказываемых ТМ на сельскохозяйственные растения. Также они углубляют понимание фундаментальных механизмов ответа растительного организма на техногенный стресс и особенностей формирования устойчивости к нему.
Список литературы
Mishra S., Bharagava N., More R.N. Heavy metal contamination: an alarming threat to environment and human health // Environmental Biotechnology: For Sustainable Future / Eds. Sobti R., Arora N., Kothari R. N.Y.: Springer, 2019. 206 p.
Demkova L., Arvay J., Bobuľska L., Tomaš J., Stanovič R., Lošak T., Harangozo L., Vollmannova A., Bystricka J., Musilova J., Jobbagy J. Accumulation and environmental risk assessment of heavy metals in soil and plants of four different ecosystems in a former polymetallic ore mining and smelting area (Slovakia) // J. Environ. Sci. Health. Part A. Toxic / Hazardous Substances Environ. Engineer. 2017. V. 52. P. 479–490.
Sharma P., Dubey R.S. Lead toxicity in plants // Braz. J. Plant. Physiol. 2005. V. 17. P. 35–52.
Журбицкий З.И. Теория и практика вегетационного метода. М.: Наука, 1968. 260 с.
Heath R.L., Packer L. Photoperoxidation in isolated chloroplasts. I. Kinetics and stoichiometry of fatty acid peroxidation // Arch. Biochem. Biophys. 1968. V. 125. № 1. P. 189–198.
Bates L.S., Waldern R.P., Teare I.D. Rapid determination of free proline for water-stress studies // Plant and Soil. 1973. V. 39. № 1. P. 205–207.
ГОСТ 12038-84. Семена сельскохозяйственных культур. Методы определения всхожести. М.: Стандартинформ, 2010. 25 с.
Паушева З.П. Практикум по цитологии растений. М.: Колос, 1974. 288 с.
Атабекова А.И., Устинова Е.И. Цитология растений. М.: Колос, 1971. 256 с.
Алов И.А. Цитофизиология и патология митоза. М.: Медицина, 1972. 264 с.
Leme D.M., Marin-Morales M.A. Alium cepa test in environmental monitoring: a review on its application // Mutation Res. 2009. V. 682. P. 71–81.
Qin R., Wang C., Chen D., Bjorn L.O., Li S. Copper-induced root growth inhibition of Allium cepa var. Agrogarum L. involves disturbances in cell division and DNA damage // Environ. Toxicol. Chem. 2015. V. 34. № 5. P. 1045–1055.
Гераськин С.А. Концепция биологического действия малых доз ионизирующего излучения на клетки // Радиац. биол. Радиоэкология. 1995. Т. 35. Вып. 5. С. 571–580.
Гераськин С.А., Дикарев В.Г., Дикарева Н.С., Удалова А.А. Влияние раздельного действия ионизирующего излучения и солей тяжелых металлов на частоту хромосомных аберраций в листовой меристеме ярового ячменя // Генетика. 1996. Т. 32. № 2. С. 272–278.
Гераськин С.А., Дикарев В.Г., Дикарева Н.С. Влияние сочетанного радиоактивного и химического (тяжелые металлы, гербицид) загрязнения на выход цитогенетических нарушений в интеркалярной меристеме ярового ячменя // Радиац. биол. Радиоэкология. 2002. Т. 42. № 4. С. 369–383.
Дикарев А.В., Дикарев В.Г., Дикарева Н.С. Влияние нитрата свинца на морфологические и цитогенетические показатели растений ярового двурядного ячменя (Hordeum vulgare L.) // Агрохимия. 2014. № 7. С. 45–52.
Дикарев А.В., Дикарев В.Г., Дикарева Н.С. Сравнительный анализ частоты цитогенетических эффектов в апикальной меристеме корешков проростков сортов ячменя (Hordeum vulgare L.), контрастных по устойчивости к свинцу // Тр. по прикл. бот., генет. и селекции. 2016. Т. 177. № 1. С. 52–68.
Дикарев В.Г., Санжарова Н.И., Гераськин С.А., Дикарев А.В., Дикарева Н.С. Влияние свинца на продуктивность, физиолого-биохимические и цитогенетические показатели ярового ячменя // Агрохимия. 2017. № 10. С. 73–83.
Дикарев В.Г., Дикарев А.В., Дикарева Н.С., Гераськин С.А. Сравнительный анализ эффективности использования интеркалярных и апикальных меристем ячменя для биоиндикации генотоксического действия свинца // Экол. генетика. 2018. Т. 16. № 3. С. 37–46.
Dikarev A.V., Dikarev V.G., Dikareva N.S. The assessment of cadmium nitrate effect on morphological and cytogenetic indeces of spring barley (Hordeum vulgare L.) seedlings // Brazil. J. Bot. 2021. V. 44. P. 43–56.
Гераськин С.А., Дикарев В.Г., Дикарева Н.С., Удалова А.А. Закономерности индукции малыми дозами ионизирующего излучения цитогенетических повреждений в корневой меристеме проростков ячменя // Радиац. биол. Радиоэкология. 1999. Т. 39. Вып. 4. С. 373–383.
Geras’kin S.A., Oudalova A.A., Kim J.K., Dikarev V.G., Dikareva N.S. Cytogenetic effect of low dose γ-radiation in Hordeum vulgare seedlings: non-linear dose–effect relationship // Radiat. Environ. Biophys. 2007. V. 46. P. 31–41.
Дополнительные материалы отсутствуют.