

Почвоведение, 2023, № 7, стр. 801-814
Подстилки и живой напочвенный покров биогеоценозов мелколиственных лесов Московской области
В. М. Телеснина a, *, О. В. Семенюк a, Л. Г. Богатырев a
a МГУ им. М.В. Ломоносова
119991 Москва, Ленинские горы, 1, Россия
* E-mail: vtelesnina@mail.ru
Поступила в редакцию 13.12.2022
После доработки 27.01.2023
Принята к публикации 28.02.2023
- EDN: VOTFEP
- DOI: 10.31857/S0032180X2260158X
Аннотация
Изучены эколого-ценотическая структура живого напочвенного покрова и структурно-функциональные особенности лесных подстилок для трех типов леса: березняка волосисто-осокового, березово-осинового леса волосисто-осокового, березняка щучкового, образующих последовательный ряд по мере увеличения гидроморфизма в пределах склона приводораздельной депрессии. Экологическая характеристика живого напочвенного покрова основана на группировке эколого-ценотических свит по А.А. Ниценко и экологических шкал Л.Г. Раменского и Х. Элленберга. Увеличение гидроморфизма сопровождается ростом эколого-ценотического разнообразия. Общий балл трофности возрастает в условиях высокого гидроморфизма в сочетании с низкими баллами отношения к кислотности по Х. Элленбергу. Напротив, максимальные баллы по этим показателям, при высоком варьировании, принадлежат березово-осиновому лесу, занимающему промежуточные позиции в ряду возрастающего гидроморфизма. Для изученных насаждений характерны деструктивные и ферментативные подстилки. По мере усиления гидроморфизма запасы подстилок увеличиваются с 400 до 1400 г/м2 с одновременным возрастанием доли детрита в подгоризонте L. Около 60% от всего запаса органического вещества, сосредоточенного в подстилках мелколиственных насаждений, приходится на легкоразлагаемые фракции. Максимальная зольность характерна для фракции детрита подгоризонта L. Параметры экологической характеристики, положенные в основу метода главных компонент, показали хорошую группировку изученных фитоценозов по степени увлажненности, особенно при использовании общих свойств подстилок (запасы, мощность, доля детрита). Выявлена целесообразность использования свойств подстилок при установлении сходства и различия изученных фитоценозов как характеристик, интегрально отражающих особенности увлажнения. Параметры живого напочвенного покрова в сопряжении с рядом структурных и функциональных особенностей лесных подстилок являются индикаторами степени гидроморфизма.
ВВЕДЕНИЕ
Подстилки мелколиственных лесов изучаются в различных целях – от вопросов их генезиса [26] до проблем мониторинга экосистем [14]. Мелколиственные леса в зонах тайги и хвойно-широколиственных лесов могут существовать как субклимаксное растительное сообщество при определенных условиях (например, повышенное увлажнение), так и в качестве стадии сукцессии. Так, роль мелколиственных лесов и характерных для них особенностей биологического круговорота признана при анализе этапов зарастания хвойных вырубок [29], при этом подчеркиваются быстрые темпы восстановлении свойств лесных почв в сочетании с коротким периодом сохранения достигнутого уровня. Подстилки мелколиственных лесов играют определенную роль в процессе постагрогенного восстановления органического вещества почв [17].
Следует отметить использование показателей опада и подстилок в мелколиственных лесах при оценке депонирования углерода [20], включая слежение за процессами деструкции. Исследуются закономерности изменения экологического состояния городских и пригородных мелколиственных лесов по сравнению с хвойными [16], в том числе на основе детального анализа лесных подстилок. Получены уникальные материалы по установлению специфики распределения корней и их запасов в подстилках [10] и почвах мелколиственных и хвойных насаждений. Показано, что гидроморфизм благоприятствует накоплению органического вещества [23], в том числе за счет травяного покрова – последний является важным фактором накопления органического вещества почвы. Для березовых насаждений южной тайги Сибири показаны особенности пространственного распределения кислотности, содержания золы и других показателей в подстилках [7, 9, 38], которые обусловлены как составом опада и живого напочвенного покрова, так и внешними факторами (глубина залегания грунтовых вод, отложения из поверхностных вод и т.д.). Зольный состав подстилки детерминирован как составом опада, так и степенью ее разложения [8].
Разнообразны зарубежные исследования мелколиственных лесов. В частности, проведена сравнительная оценка потоков углерода в системе подстилок сосново-еловых и березовых насаждений [39]. Ряд работ посвящен связыванию углерода в березовых лесах в зависимости от атмосферного увлажнения [33, 37]. Осиновые насаждения и накапливаемое ими органическое вещество почв и подстилок имеют особенности в связи с составом древесного опада и специфическим микроклиматом, который создает осина (накопление влаги подстилкой, относительно повышенное богатство почвы элементами питания при высокой затененности). Осиновые насаждения предложено занести в глобальный природоохранный контекст, так они могут служить моделью для сохранения биоразнообразия [36]. Показано влияние дополнительного внесения азота и серы на процессы разложения в осиновом лесу [40]. Проведены исследования по оценке дыхания в зимний период в полевых и лабораторных условиях для подстилок осинового леса [30, 35].
Подстилки мелколиственных лесов, особенно заболоченных, на сегодняшний день изучены недостаточно [9], что связано с ограниченным распространением подобных экосистем. Вместе с тем очень важно понимание влияния степени гидроморфизма, а также состава мелколиственного древостоя на аккумуляцию подстилкой органического углерода. Важным индикатором характера увлажнения является живой напочвенный покров, который быстрее реагирует на локальные экологические условия и их изменения, нежели древостой [12, 13, 28]. В частности, в заболоченных и болотных экосистемах определенные виды живого напочвенного покрова и их сочетания соответствуют эвтрофному, олиготрофному или мезотрофному типам торфа [1].
Показатели живого напочвенного покрова и подстилок имеют существенную информационную значимость для ряда характеристик лесной экосистемы.
Цель работы – изучить индикационную роль живого напочвенного покрова и лесных подстилок в определении степени гидроморфизма и интенсивности биологического круговорота применительно к мелколиственным лесам. В задачи исследования входило:
1) определение влияния увлажнения на параметры живого напочвенного покрова (биомассу, флористический состав и эколого-ценотические характеристики),
2) определение структурных и функциональных параметров подстилок в насаждениях, различающихся по степени увлажненности и составу древостоя.
ОБЪЕКТЫ И МЕТОДЫ
Исследования проводили на территории Солнечногорского района Московской области в окрестностях учебно-опытного почвенно-экологического центра “Чашниково”. Объектами послужили три мелколиственных биогеоценоза, расположенных на пологом склоне, переходящем в приводораздельную депрессию и образующих единую почвенно-геохимическую катену [25]. В пределах катены вниз по склону последовательно увеличивается гидроморфизм местообитания и влажность почв, изменяется характер живого напочвенного покрова, что связано с повышением уровня залегания почвенно-грунтовых вод. Почвообразующие породы представлены покровными суглинками, подстилаемыми мореной.
Березняк волосисто-осоковый расположен в пределах краевой верхней части приводораздельной депрессии, прилегающей к опытным полям агробиостанции. Растительность представлена березовым лесом (древостой 10Б), в напочвенном покрове которого преобладают осока волосистая (Carex pilosa) – 80%, зеленчук желтый (Galeobdolon luteum) и копытень европейский (Asarum europaeum). Почва – дерново-подзолисто-глеевая [11] (Albic Gleyic Retisol (Loamic)). Профиль состоит из следующих горизонтов: О(0–3 см)–AY(3–14)–AEL(14–20)–ELg(20–34)–BELg(34–55)–BTg(55–87)–BCg. Легкий суглинок на глубине горизонта ELg становится опесчаненным, при переходе к BELg и глубже – средним суглинком. Почвенно-грунтовые воды вскрываются на глубине более 100–120 см. Следует отметить, что данный фитоценоз наиболее приближен к находящемуся выше по склону распаханному полю, которое может являться источником дополнительного поступления мелкозема с делювиальным стоком.
Березово-осиновый волосисто-осоковый лес (древостой 8О2Б) расположен в середине склона, примерно в 75–100 м от предыдущего фитоценоза. В напочвенном покрове преобладает осока волосистая (22%), но появляются такие виды, как щитовник мужской (Dryopreris filix-mass), хвощ лесной (Equisetum sylvaticum) и вербейник обыкновенный (Lysimachia vulgaris). Почва – перегнойно-подзолисто-глеевая (Albic Gleyic Retisol (Loamic)). Строение профиля: О(0–5 см)–H(5–13)–AEL(13–17)–Elg(17–36)–BELg(36–53)–BTg(53–81)–BCg. Горизонты AEL и EL – легкосуглинистые, нижележащие горизонты – среднесуглинистые. Почвенно-грунтовые воды вскрываются на глубине 80–110 см.
Березняк щучковый расположен недалеко от центра депрессии рельефа в нижней части катены, примерно в 50–70 м от березово-осинового леса. Растительность представлена березовым лесом (древостой 10Б), в напочвенном покрове преобладают виды, соответствующие условиям повышенного увлажнения: щучка дернистая (Deschampsia caespitosa) – 50%, ситник тонкий (Juncus tennius) – 5%, таволга вязолистная (Filipendula ulmaria) – 10%. Кроме того, фрагментарно развит моховой ярус, образованный Pleurozium shreberii. Почва – торфяно-перегнойно-подзолисто-глеевая (Albic Gleyic Histic Retisol (Loamic)). Строение профиля: О(0–8 см)–T(8–13)–H(13–25)–ELg(25–39)–BELg(39–50)–BTg. Легкий суглинок в горизонте ELg ниже сменяется средним суглинком. Почвенно-грунтовые воды вскрываются на глубине 40–60 см.
Во всех фитоценозах на площади 10 × 10 м провели геоботанические описания, а также отбор укосов живого напочвенного покрова с площадок 50 × 50 см в 5-кратной повторности (точки отбора расположены случайно примерно в 5 м друг от друга) с последующим разбором на виды. Запасы биомассы живого напочвенного покрова рассчитывали на абсолютно сухое вещество (высушенное при 105°С) в г/м2. Для экологических характеристик напочвенного покрова использовали классификацию эколого-ценотических свит по А.А. Ниценко [18], а также экологические шкалы Л.Г. Раменского [22], Э. Ландольта [32] и Х. Элленберга [31]. С помощью шкал Л.Г. Раменского для живого напочвенного покрова каждого из укосов рассчитывали средневзвешенные баллы влажности и трофности (средневзвешенная середина интервала). В соответствии со шкалой отношения растений к кислотности почвы (по Х. Элленбергу) рассчитывали средневзвешенный балл отношения к кислотности. В полевых условиях проводили морфологическое описание подстилок и определение мощности каждого их подгоризонта. Классификационную принадлежность подстилок определяли по Л.Г. Богатыреву [3]. В соответствии с этой классификацией, деструктивные маломощные подстилки морфологически состоят из опада прошлых лет (единственный подгоризонт L), что свидетельствует о высокой скорости трансформации растительного опада. При снижении скорости разложения опада формируются подстилки более сложного строения ферментативные (L–F), которые кроме подгоризонта L включат ферментативный (F) подгоризонт, характеризующийся средней степенью разложения органического вещества. Образцы лесных подстилок отбирали в 9-кратной повторности с площадок 25 × 25 см (все подгоризонты отдельно). Повторности в пределах участка расположены случайно – примерно 1 на 10 м2, поскольку в приствольных и подкроновых пространствах существенных различий между подстилками в данном случае нет. Запасы подстилок и их компонентов рассчитывали на абсолютно сухое вещество в г/м2. Подоризонт L разбирали на отдельные компоненты: листья, ветошь (опад трав), шишки, ветки и т.д. К детриту относили компоненты L, происхождение которых установить не представлялось возможным. Помимо запасов и компонентного состава подстилок, рассчитывали долю легкоразлагаемых компонентов (листьев и ветоши) и детрита в подгоризонте L, а также соотношение запасов подгоризонтов L и F. Зольность подстилок и их компонентов определяли путем сжигания при температуре 450°С. Рассчитывали показатели средневзвешенной зольности подгоризонта L, запасов зольных элементов в L, запасы зольных элементов в легкоразлагаемых компонентах (листья и ветошь – под последней понимали опад трав) и их отношение к общим запасам золы в подгоризонте L. Определяли рН водной суспензии во всех подгоризонтах подстилок. Обработку полученных результатов осуществляли с помощью программ Exсel и Statistica.
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
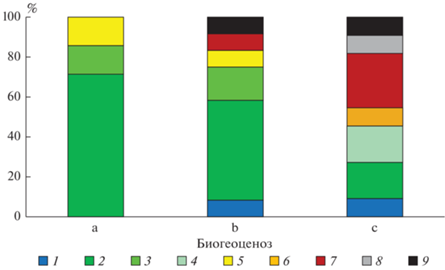
Растительность. Живой напочвенный покров изученных биогеоценозов существенно различается. Для березняка волосисто-осокового выявлено наименьшее разнообразие эколого-ценотических свит (рис. 1), среди которых преобладает неморальная теневая (осока волосистая – Carex pilosa), а также встречаются виды других неморальных свит. В березняке щучковом, где увлажнение существенно больше, число свит увеличивается вдвое, в том числе за счет видов торфяно-луговой (щучка дернистая Deschampsia caespitosa), черноольховой (таволга вязолистная Filipendula ulmaria) и болотно-кустарничковой (вербейник обыкновенный Lysimachia vulgaris) свит. Спецификой эколого-ценотической структуры живого напочвенного покрова березово-осинового леса является преобладание неморальных и еловых черничных свит, характерных для дренированных почв, в сочетании с наличием видов болотно-кустарничковой группы, которая служит индикатором повышенного увлажнения.
Рис. 1.
Соотношение видов разных эколого-ценотических свит в живом напочвенном покрове: 1 – еловая черничная, 2 – неморальная теневая, 3 – неморальная опушечно-полянная, 4 – неморальная высокотравная полянная, 5 – полунеморальная, 6 – черноольховая, 7 – болотная кустарничковая, 8 – торфяно-луговая, 9 – эвритопы. a – березняк волосисто-осоковый; b – березово-осиновый волосисто-осоковый; c – березняк щучковый.
Надземная биомасса живого напочвенного покрова значимо повышена в березняке щучковом по сравнению с другими фитоценозами (рис. 2) – за счет доминирования высокопродуктивных видов, таких как щучка дернистая и камыш лесной (Scirpus sylvaticus). В то же время именно для заболоченного березняка выявлено максимальное варьирование биомассы живого напочвенного покрова в пространстве, что связано с увеличением его видового и эколого-ценотического разнообразия. Для этого фитоценоза установлен максимальный средневзвешенный балл увлажнения по Л.Г. Раменскому.
Рис. 2.
Количественные характеристики живого напочвенного покрова изученных фитоценозов: a – биомасса живого напочвенного покрова, г/м2; b – средневзвешенный балл увлажнения по Раменскому; c – средний балл трофности по Раменскому; d – средневзвешенный балл отношения к кислотности по Элленбергу. 1 – березняк волосисто-осоковый, 2 – березово-осиновый волосисто-осоковый, 3 – березняк щучковый. I – среднее, II – стандартная ошибка, III – стандартное отклонение.
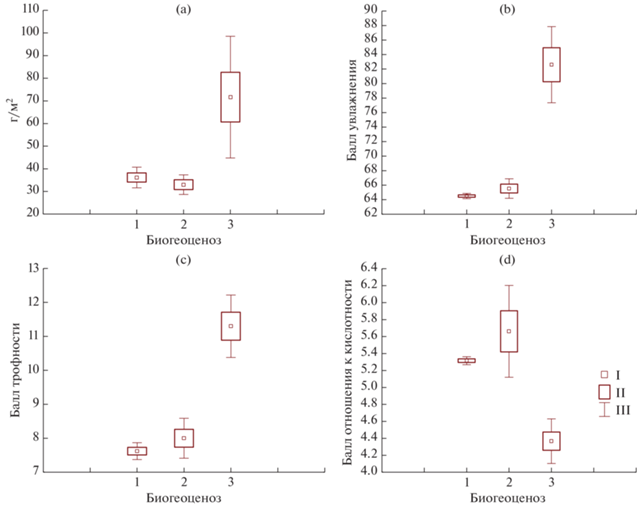
Балл трофности возрастает вниз по склону, наиболее значимо увеличиваясь в березняке щучковом. В то же время для живого напочвенного покрова березняка щучкового выявлены самые низкие средневзвешенные баллы отношения к кислотности по Х. Элленбергу, что говорит о преобладании ацидофилов. Максимальный балл отношения к кислотности соответствует березово-осиновому лесу, где видов-ацидофилов гораздо меньше. Как видно, трофность является сложной категорией, которая не связана напрямую с отношением к кислотности и другими отдельно взятыми показателями, о чем неоднократно упоминалось в литературе [2, 34]. Вероятно, трофность в данном случае связана с относительной обогащенностью почвенно-грунтовых вод различными макроэлементами, в том числе кальцием и магнием, а также с возможной латеральной миграцией вод вниз по склону. Действительно, ранее для данной катены установлено увеличение содержания кальция и магния в почвенных вытяжках с <1 до 20 мг/л вниз по склону [4], что детерминировано в том числе составом почвенно-грунтовых вод, глубина залегания которых вниз по склону уменьшается.
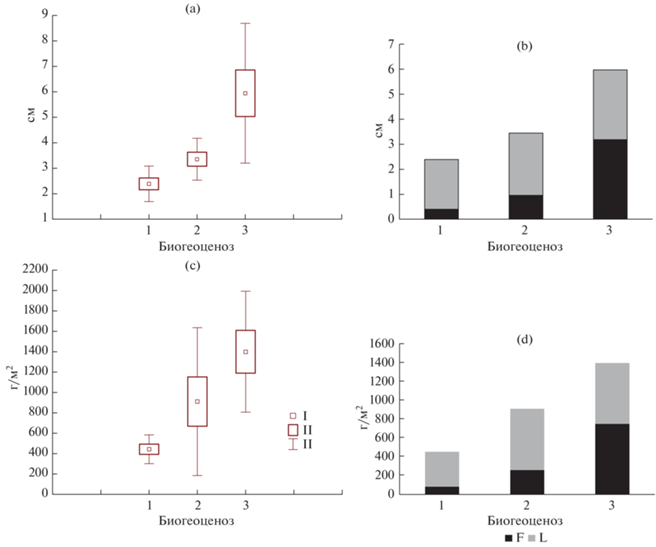
Подстилки. Подстилки мелколиственных насаждений представлены деструктивным и ферментативным типами с преобладанием ферментативного типа (>70%). По мере увеличения степени увлажнения биогеоценозов доля ферментативных подстилок увеличивается, и в березняке щучковом встречаются только ферментативные подстилки. В ряду березняк волосисто-осоковый – березово-осиновый волосисто-осоковый – березняк щучковый значимо увеличиваются мощность и запасы подстилок, достигая максимума по мощности (6 см) и запасам (1400 г/м2) в последнем биогеоценозе (рис. 3). По мере увеличения увлажнения в общих запасах подстилки существенно возрастает доля запасов верхнего деструктивного подгоризонта L c 20 до 60%.
Рис. 3.
Мощность подстилки, см: a – общая, b – по подгоризонтам (L и F). Запасы подстилки, г/м2: c – общие, d – по подгоризонтам. 1 – березняк волосисто-осоковый, 2 – березово-осиновый волосисто-осоковый, 3 – березняк щучковый. I – среднее, II – стандартная ошибка, III – стандартное отклонение.
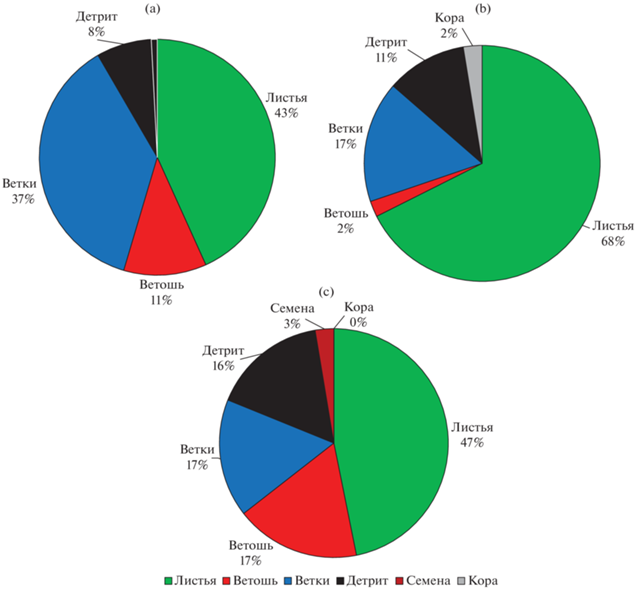
Несмотря на то, что для подстилок всех насаждений преобладающими фракциями являются листья (40–60%) и ветки (20–40%), именно в подстилках березово-осинового леса наблюдается максимальная доля фракции листьев (63.6%), а в березняке щучковом – максимальная доля фракции ветоши, вероятно, из-за высокой продуктивности травяного яруса. Наличие значительного объема легкоразлагаемых компонентов в подстилках березово-осинового волосисто-осокового леса связано с большим объемом поступающего листового опада осины, тогда как в подстилках березняка щучкового – с повышенным увлажнением, которое создает неблагоприятные условия для функционирования микроорганизмов и снижает скорость вовлечения органического вещества и зольных элементов в биологический круговорот. По мере увеличения увлажнения, в 2 раза возрастает долевое участие фракции детрита в подгоризонте L (рис. 4), запасы его при этом увеличиваются в 3 раза.
Рис. 4.
Фракционный состав подгоризонта L, %: a – березняк волосисто-осоковый, b – березово-осиновый волосисто-осоковый, c – березняк щучковый.
Определение содержания зольных элементов во фракциях подстилки березняка щучкового (табл. 1) показало, что максимальной зольностью характеризуются детрит (15%) и ветошь (22%). Зольность листьев (9%) ниже зольности детрита, тогда как зольность веток еще ниже (4%), что соответствует литературным данным [24, 27]. Зольность всех фракций в подстилках березняка волосисто-осокового больше, чем аналогичных фракций в березняке щучковом, что, прежде всего, относится к листьям, семенам и детриту, а также содержанию золы в подгоризонте F (в 2–3 раза). Для подстилок березово-осинового леса почти все эти характеристики в 1.4–2 раза превышают значения соответствующих показателей для березняка щучкового. Значительные различия между изученными биогеоценозами отмечаются и по другим показателям функционирования подстилок (табл. 2). По мере усиления гидроморфизма величина соотношения мощности подгоризонтов L/F снижается в 5 раз, тогда как запасы детрита увеличиваются в 3 раза.
Таблица 1.
Зольность компонентов подстилок, % (± ошибка среднего)
| Горизонт | Компонент | Березняк волосисто-осоковый | Березово-осиновый волосисто-осоковый | Березняк щучковый |
|---|---|---|---|---|
| L | Листья | 19 ± 2 | 17 ± 1 | 9 ± 3 |
| Ветви | 7 ± 1 | 12 ± 3 | 4 ± 1 | |
| Ветошь | 30 ± 6 | 23 ± 2 | 25 ± 8 | |
| Семена | 13 ± 3 | 8 ± 2 | 4 ± 0.4 | |
| Детрит | 28 ± 3 | 36 ± 5 | 15 ± 2 | |
| F | 41 ± 10 | 43 ± 5 | 22 ± 5 |
Таблица 2.
Показатели функционирования подстилок
| Параметр | Березняк волосисто-осоковый | Березово-осиновый волосисто-осоковый | Березняк щучковый | |
|---|---|---|---|---|
| Соотношение L/F | 4.5 | 2.5 | 0.9 | |
| Легкоразлагаемые компоненты | Запасы, г/м2 | 201 ± 8 | 479 ± 22 | 417 ± 16 |
| Доля, % | 55.3 | 62.7 | 60.3 | |
| Детрит | Запасы, г/м2 | 28 ± 5 | 71 ± 8 | 105 ± 8 |
| Доля, % | 7.5 | 14.9 | 23.7 | |
| Тип подстилки | % деструктивных | 37 | 22 | 0 |
| % ферментативных | 63 | 78 | 100 | |
| Средневзвешенная зольность горизонта L, % | 14 ± 2 | 18 ± 2 | 9 ± 1 | |
| Запас зольных элементов в подгоризонте L, г/м2 | 45 ± 5 | 98 ± 23 | 72 ± 8 | |
| Общие запасы зольных элементов в подстилке, г/м2 | 62.2 | 231.8 | 230.0 | |
| Запасы зольных элементов в составе легкоразлагаемых компонентов, г/м2 | 30 ± 2 | 94 ± 25 | 50 ± 8 | |
| Доля зольных элементов легкоразлагаемых фракций в общей сумме зольных элементов подгоризонта L, % | 66.5 | 97.0 | 69.4 | |
Запас легкоразлагаемых компонентов максимален в березово-осиновом волосисто-осоковом лесу и составляет 479 г/м2, что сопоставимо с запасом в подстилках березняка щучкового и в 2 раза превышает запас легкоразлагаемых компонентов в подстилках березняка волосисто-осокового. Аналогичны закономерности для запасов золы подстилки в общем и золы подгоризонта L в частности. При этом средневзвешенная зольность подгоризонта L в подстилках березняка волосисто-осокового в 1.5 раза выше по сравнению с березняком щучковым и несколько ниже, чем в березово-осиновом. Таким образом, подстилки березово-осинового леса отличаются повышенным запасом фракции листьев. Согласно литературным данным, зольность листьев осины больше, нежели зольность листьев березы [20, 21], что обусловливает отличие подстилок березово-осинового леса от других по перечисленным показателям. Необходимо отметить, что повышенные значения зольности компонентов подстилки березняка волосисто-осокового, который наиболее близко расположен к сельскохозяйственным угодьям выше по склону, показывают, что для подстилок этот показатель может отражать влияние механической миграции, проявляющейся в виде ветрового переноса и поверхностного стока твердых частиц с полей.
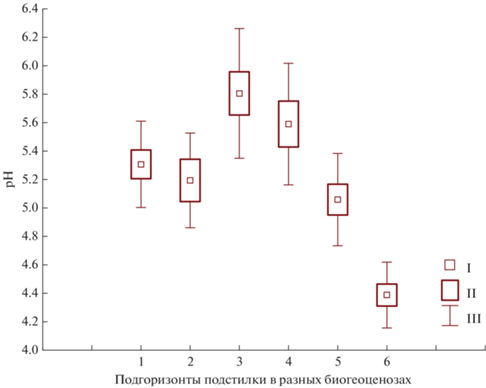
Кислотность подстилок имеет тенденцию к увеличению от L к F (рис. 5), но значимые различия между L и F выявлены только для подстилок березняка щучкового, поскольку подстилки этого фитоценоза как наиболее увлажненного имеют более сложное строение. Именно подгоризонт F, который в отличие от L связан в большей степени с факторами разложения органического вещества, нежели с химическим составом опада, реагирует повышенной кислотностью на гидрологический режим как важный фактор кислотных свойств подстилки [9]. Следует отметить, что распределение показателей рН подстилок, особенно ферментативных подгоризонтов, адекватно данным, полученным для баллов отношения растений травяного яруса к кислотности – наибольшая кислотность подстилок соответствует наименьшему баллу отношения к кислотности видов живого напочвенного покрова. Для подстилок березово-осинового леса отмечено наибольшее значение рН (5.8), что значимо выше величины реакции среды подстилок березняков (рН 5.2). Возможно, последнее связано с содержанием в осиновых листьях кальция, которое больше, нежели в березовых [20].
Рис. 5.
Величина рН подстилок: 1 – березняк волосисто-осоковый, L, 2 – березняк волосисто-осоковый, F, 3 – березово-осиновый волосисто-осоковый, L, 4 – березово-осиновый волосисто-осоковый, F, 5 – березняк щучковый, L, 6 – березняк щучковый, F. I – среднее, II – стандартная ошибка, III – стандартное отклонение.
Многие из определенных параметров как живого напочвенного покрова, так и подстилок, отличаются для изученных фитоценозов не только по абсолютным величинам, но и по степени пространственного варьирования. Так, существенно увеличена по сравнению с другими фитоценозами биомасса живого напочвенного покрова в березняке щучковом, а также ее варьирование внутри фитоценоза – последнее связано с увеличением флористического и эколого-ценотического разнообразия травянистых растений, имеющих разную продуктивность. Общие запасы подстилки сильно варьируют в березово-осиновом лесу, возможно, это обусловлено повышенным разнообразием локальных экологических ниш фитоценоза, занимающего промежуточное положения по фактору увлажненности.
Для того, чтобы выявить степень влияния различных факторов на особенности живого напочвенного покрова, а также показатели структуры и функционирования лесных подстилок, все полученные данные были проанализированы методом главных компонент. Проведены три серии анализа по следующим группам свойств (табл. 3): характеристики живого напочвенного покрова; показатели подстилок, связанные непосредственно с растительным опадом; показатели подстилки, связанные со степенью увлажнения.
Таблица 3.
Факторные координаты переменных на основе корреляций
| Переменная | Фактор 1 | Фактор 2 |
|---|---|---|
| Анализ с учетом характеристик живого напочвенного покрова | ||
| Биомасса | 0.794429 | –0.218523 |
| Доля видов водно-болотных свит | 0.929238 | –0.061177 |
| Балл увлажнения (по Раменскому) | 0.736724 | –0.264188 |
| Балл трофности (по Раменскому) | 0.593499 | –0.544928 |
| Доля влаголюбивых видов (по Элленбергу) | 0.825666 | –0.012949 |
| Доля видов, соответствующих органогенным почвам (по Ландольту) | 0.939828 | –0.111051 |
| Доля видов, соответствующих нейтральным почвам (по Элленбергу) | –0.639263 | –0.739402 |
| Доля видов-нитрофилов (по Элленбергу) | –0.455425 | –0.858284 |
| Анализ с учетом показателей биологического круговорота | ||
| Запасы фракции листьев в L | –0.487413 | 0.839442 |
| Запасы фракции ветоши в L | 0.799649 | 0.552158 |
| Доля фракции листьев в L | –0.676651 | 0.436721 |
| Доля фракции ветоши в L | 0.870179 | 0.237558 |
| Зольность фракции листьев | –0.476820 | –0.173514 |
| Запас золы фракции листьев | –0.640830 | 0.692320 |
| Запас золы фракции ветоши | 0.788301 | 0.529413 |
| Анализ с учетом показателей подстилки, связанных со степенью увлажнения | ||
| Мощность подстилки | –0.751349 | –0.094330 |
| Запасы общие | –0.851117 | 0.335517 |
| Запасы F | –0.894910 | 0.234018 |
| Запасы детрита | –0.319379 | –0.910902 |
| Доля детрита в L | –0.139345 | –0.955845 |
Анализ данных по живому напочвенному покрову (табл. 3) показал, что фактор 1 связан с долей видов, соответствующих высокой степени увлажненности и предпочитающих органогенные почвы (коэффициенты корреляции 0.82–0.94). Поэтому точки 11–15 (березняк щучковый) образуют выраженную отдельную группу (рис. 6а). Березняк волосисто-осоковый отчетливо отделен от общего массива, поскольку характеризуется чрезвычайно низким разнообразием живого напочвенного покрова, в котором полностью отсутствуют влаголюбивые виды.
Рис. 6.
Результаты анализа данных методом главных компонент, выполненных с учетом: a – характеристик живого напочвенного покрова; b – показателей биологического круговорота; c – показателей подстилки, связанных со степенью увлажнения. Расположение объектов исследования в плоскости двух первых компонент. a: 1–5 – березняк волосисто-осоковый, 6–10 – березово-осиновый волосисто-осоковый, 11–15 – березняк щучковый. b, c: 1–9 – березняк волосисто-осоковый, 10–18 – березово-осиновый волосисто-осоковый, 19–27 – березняк щучковый.
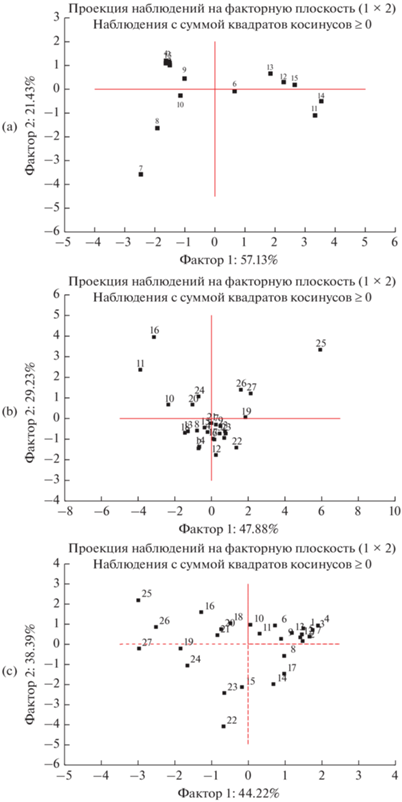
Результаты анализа по свойствам подстилок, непосредственно связанных с увлажнением, показывают, что фактор 1, отрицательно коррелирующий с запасами подстилки в целом и подгоризонта F (–0.85 и –0.89 соответственно), почти полностью отделяет подстилки березняка щучкового как наиболее увлажненного от остальной совокупности. Повторности, которые соответствуют березово-осиновому лесу, не так обособлены на графике. Видимо, промежуточное положение по гидроморфизму обусловливает относительно высокое разнообразие локальных экологических ниш. Фактор 2 коррелирует с долей и запасами детрита и отделяет группу повторностей, соответствующих наиболее дренированному березняку волосисто-осоковому.
Анализ данных по свойствам подстилок, связанным с опадом, показал следующее. Фактор 1, связанный с запасами и долей ветоши (коэффициент 0.8–0.85), отделяет почти все точки, соответствующие березово-осиновому лесу, от остальных точек. Действительно, именно в этом лесу выявлена очень низкая биомасса живого напочвенного покрова и, как следствие, низкий запас ветоши. Следовательно, надземный опад трав не влияет на емкость и интенсивность биологического круговорота. Доля листьев в подгоризонте L, а также запасы зольных элементов в них существенно отличаются в большую сторону от остальных фитоценозов. Точки, соответствующие березняку щучковому, также составляют отчетливую группу, поскольку запасы биомассы и, соответственно, ветоши здесь, напротив, максимальны из трех фитоценозов.
В условиях повышенной влажности снижается активность функционирования микроорганизмов и скорость разложения органического вещества, что способствует его накоплению в форме слаборазложившихся остатков (фракция детрита в подгоризонте L). Уменьшение соотношения мощности и запасов подгоризонтов L/F также свидетельствует о накоплении органического вещества в разной степени разложения, но уже в ферментативном F. Влияние гидроморфизма на скорость разложения органического вещества и, как следствие, интенсивность круговорота, особенно показательно при сравнении свойств подстилок, соответствующих фитоценозам с одинаковым эдификаторам: березняков волосисто-осокового и щучкового.
Влияние дерева-эдификатора на свойства подстилок проявляется, прежде всего, в соотношении фракции листьев и веток. Для подстилки березово-осинового леса отмечена максимальная зольность подгоризонтов L и F, а также запасов зольных элементов, что связано с высокой обогащенностью осинового опада зольными элементами [6, 15, 26].
Влияние живого напочвенного покрова на фракционный состав наиболее ярко проявляется в березняке щучковом, где значительная фитомасса напочвенного покрова соответствует максимальному долевому участию ветоши в фракционном составе подстилки данного фитоценоза. Живой напочвенный покров вносит определенный вклад в накопление подстилкой зольных элементов, в формирование определенного соотношения углерода и азота и, как следствие, на скорость преобразования органического вещества [19].
В данном случае имеют место несколько факторов, определяющих структурные и функциональные особенности, а также некоторые химические свойства подстилок мелколиственных лесов:
1) степень гидроморфизма, которая зависит от уровня залегания грунтовых вод, которые могут быть источником дополнительного поступления элементов питания,
2) состав древостоя – в данном случае соотношение березы и осины. Строение кроны, в свою очередь, детерминирует развитие травяного яруса благодаря фактору затенения. Живой напочвенный покров, хотя и мало участвует в биологическом круговороте по сравнению с опадом древостоя, служит четким индикатором условий увлажнения и, как следствие, особенностей накопления наземного детрита в виде подстилок,
3) степень удаленности от источника поступления минеральных частиц на поверхность лесной подстилки (распаханное поле вверх по склону).
В березняке волосисто-осоковом минимально выражен гидроморфизм, что находит отражение в малой мощности и запасах подстилок, а также относительно низком запасе детрита. Высокая зольность многих компонентов подстилки по сравнению с зольностью аналогичных компонентов в другом березняке, вероятно, обусловлена именно привносом минеральных компонентов.
Березово-осиновый волосисто-осоковый лес характеризуется не только повышенным поступлением зольных элементов с опадом листвы осины, но и более высокой степенью гидроморфизма. Однако в данном случае почвенно-грунтовые воды расположены глубоко (80–110 см), чтобы не вызывать постоянный застой влаги, который существенно снижал бы интенсивность разложения органического вещества. Березово-осиновый фитоценоз, занимая промежуточное положение по увлажненности, существенно отличается от остальных фитоценозов по следующим признакам, не связанным со степенью гидроморфизма: 1) эдификатор (осина, отличающаяся листьями с высокой зольностью); 2) крайне низкая роль живого напочвенного покрова в круговороте (по причине затенения, создаваемого осиной). Осиновый опад характеризуется соотношением C/N 118, тогда как березовый – 76 [5], что создает неоднозначную картину интенсивности разложения осинового опада, несмотря на повышенную зольность последнего, а также повышенное содержание кальция и калия по сравнению с березовым листовым опадом. Эти различия находят существенное отражение в свойствах подстилок, таких как запасы легко разлагаемых фракций, зольность компонентов и запасы зольных элементов в подстилке и ее фракциях.
Наконец, березняк щучковый наиболее увлажнен, а также максимально удален от источника поверхностного поступления минеральных компонентов. При постоянном переувлажнении почв разложение органического вещества замедлено, что находит отражение на таких показателях, как мощность, запасы подстилки, запасы детрита, значения которых существенно превышают аналогичные значения для менее увлажненного березняка. При этом зольность почти всех компонентов подстилки березняка щучкового ниже, чем в подстилках березняка волосисто-осокового, и подстилки первого накапливают зольные элементы исключительно за счет своей массы, а не за счет высокой зольности. Возможно, поступление дополнительных элементов минерального питания с почвенно-грунтовыми водами несколько увеличивает скорость разложения органического вещества, поскольку при такой степени увлажненности ферментативные подстилки несколько нетипичны – обычно при аналогичных условиях увлажнения развиты гумифицированные и(или) перегнойные подстилки [7].
ЗАКЛЮЧЕНИЕ
Живой напочвенный покров является индикатором степени увлажнения, при возрастании последней увеличивается не только доля влаголюбивых видов, но и доля ацидофилов. Биомасса живого напочвенного покрова максимальна для наиболее увлажненных насаждений. Смена деструктивных типов подстилок на ферментативные с одновременным повышением запасов подстилок и доли детрита в подгоризонте L в ряду изученных типов леса является показателем снижения скорости разложения органического вещества при увеличении степени увлажнения. Подстилки осиновых насаждений по сравнению с подстилками березовых содержат в подгоризонте L больше листовой фракции, а также накапливают больше зольных элементов почти во всех фракциях. Установлена взаимосвязь живого напочвенного покрова и подстилок как сопряженных индикаторов степени гидроморфности почв. Показано, что полученные характеристики по степени возрастания информативности для установления уровня различия экосистем заболоченных мелколиственных лесов окрестностей Чашникова можно выстроить в ряд: биомасса живого напочвенного покрова и состав древесного опада – экологические параметры живого напочвенного покрова – тип и характеристика подстилок.
Список литературы
Аветов Н.А., Кузнецов О.Л., Шишконакова Е.А. Почвы олиго-мезотрофных и мезотрофных болот бореального пояса Западной Сибири: возможности геоботанической диагностики в рамках типа торфяных мезотрофных почв // Почвоведение. 2021. № 5.
Аветов Н.А., Шишконакова Е.А. Некоторые аспекты систематики и диагностики торфяных почв бореальных болот // Почвоведение. 2019. № 8. С. 901–909.
Богатырев Л.Г. О классификации лесных подстилок // Почвоведение. 1990. № 3. С. 118–127.
Богатырев Л.Г., Бенедиктова А.И., Телеснина В.М., Карпухин М.М., Жилин Н.И., Земсков Ф.И., Демин В.В. Водные вытяжки как критерий оценки геохимической обстановки в условиях монолитной почвенно-геохимической катены в пределах верхнего течения Клязьмы // Вестник Моск. ун-та. 2020. № 4. С. 17–26.
Ведрова Э.Ф., Решетникова Т.В. Масса подстилки и интенсивность ее разложения в 40-летних культурах основных лесообразующих пород Сибири // Лесоведение. 2014. № 1. С. 42–50.
Демаков Ю.П., Исаев А.В., Шарафутдинов Р.Н. Роль лесной подстилки в борах Марийского Заволжья и вариабельность ее параметров // Науч. Тр. гос. природного заповедника “Большая Кокшага”. 2018. № 8. С. 15–43.
Ефремова Т.Т., Аврова А.Ф., Ефремов С.П., Мелентьева Н.В. Стадийность трансформации органического вещества подстилок болотных березняков // Почвоведение. 2009. № 10. С. 1203–1212.
Ефремова Т.Т., Ефремов С.П., Аврова А.Ф. Зольный состав морфометрических фракций как показатель стадий преобразования подстилок (на примере болотных березняков) // Почвоведение. 2022. № 11. С. 1351–1365. https://doi.org/10.31857/S0032180X2211003X
Ефремова Т.Т., Секретенко О.П., Аврова А.Ф., Ефремов С.П. Пространственная структура кислотных свойств подстилки в сукцессионном ряду болотных березняков // Известия РАН. Сер. биологическая. 2013. № 5. С. 624–636.
Карпечко А.Ю., Туюнен А.В., Медведева М.В., М-ошкина Е.В., Дубровина И.А., Геникова Н.В., Сидорова В.А., Мамай А.В., Толстогузов О.В., Кулакова Л.М. Масса тонких корней в почвах лесных сообществ на постагрогенных землях в условиях средней тайги (на примере республики Карелия) // Растительные ресурсы. 2021. Т. 57. Вып. 2. С. 145–157.
Классификация и диагностика почв России. Смоленск: Ойкумена, 2004.
Клещева Е.А. Использование экологических шкал для индикации современного состояния лесных сообществ (на примере сосновых лесов) // Экология. 2007. № 2. С. 104–110.
Копцик Г.Н., Багдасарова Т.В., Горленко О.В. Взаимосвязи видового разнообразия растений и свойств почв в экосистемах южной тайги // Бюл. МОИП. отд. биол. 2001. Т. 106. Вып. 2. С. 31–38.
Королькова Е.О. Мониторинг рекреационного использования особо охраняемых природных территорий на примере Полистовского государственного природного заповедника // Социально-экологические технологии. 2015. № 1–2. С. 30–39.
Кузнецов М.А. Влияние условий разложения и состава опада на характеристики и запас подстилки в среднетаежном чернично-сфагновом ельнике // Лесоведение. 2010. № 6. С. 54–60.
Мошкина Е.В., Мамай А.В. Оценка плодородия и экологического состояния автоморфных почв городских и пригородных лесов // Вестник современной науки. 2016. № 10. С. 31–37.
Назарюк В.М., Калимуллина Ф.Р. Роль природных экосистем в восстановлении плодородия выпаханных почв Западной Сибири // Проблемы агрохимии и экологии. 2017. № 1. С. 43–50.
Ниценко А.А. Об изучении экологической структуры растительного покрова // Бот. журн. 1969. Т. 54. № 7. С. 1002–1014.
Припутина И.В., Фролова Г.Г., Шанин В.Н., Мякшина Т.Н., Грабарник П.Я. Распределение органического вещества и азота в дерново-подбурах приокско-террасного заповедника и его связь со структурой лесных фитоценозов // Почвоведение. 2020. № 8. С. 921–933.
Пристова Т.А. Биологический круговорот веществ во вторичном лиственно-хвойном насаждении средней тайги // Вестник Института Коми научного центр РАН. 2006. № 8(106). С. 7–12.
Пристова Т.А. Компоненты углеродного цикла в лиственно-хвойном насаждении средней тайги // Лесоведение. 2010. № 6. С. 12–19.
Раменский Л.Г., Цаценкин И.А., Чижиков О.Н., Антипов Н.А. Экологическая оценка кормовых угодий по растительному покрову. М.: Сельхозгиз, 1956. 472 с.
Ревина О.А., Ревин А.Г. Биогеохимические особенности элементарных ландшафтов памятника природы регионального значения “Красный бор” // Природа и общество: в поисках гармонии. 2019. №. 5. С. 217–227.
Семенюк О.В., Телеснина В.М., Богатырев Л.Г., Бенедиктова А.И., Кузнецова Я.Д. Оценка внутрибиогеоценозной изменчивости лесных подстилок и травяно-кустарничковой растительности в еловых насаждениях // Почвоведение. 2020. № 1. С. 31–43.
Стома Г.В., Богатырев Л.Г., Макаров М.И., Манахов Д.В. Летняя практика по почвоведению: Уч-метод. Пособие для студентов 1 курса факультета почвоведения МГУ. М.: МАКС-Пресс, 2017. 156 с.
Тарасов П.А., Тарасова А.В., Иванов В.А. Основные характеристики лесной подстилки производных мелколиственных насаждений // Вестник КрасГУ. 2015. № 2. С. 197–200.
Телеснина В.М., Богатырев Л.Г., Бенедиктова А.И., Земсков Ф.И., Маслов М.Н. Динамика поступления растительного опада и некоторых свойств лесных подстилок при постагрогенном лесовосстановлении в условиях южной тайги // Вестник Моск. ун-та. Сер. 17, почвоведение. 2019. № 4. С. 3–10.
Телеснина В.М., Семенюк О.В., Богатырев Л.Г. Свойства лесных подстилок во взаимосвязи с напочвенным покровом в лесных экосистемах Подмосковья (на примере УОПЭЦ “Чашниково”) // Вестник Моск. ун-та. Сер. 17, почвоведение. 2017. № 4. С. 11–20.
Трефилова О.В., Ефимов Д.Ю. Изменение растительного покрова и почв при естественном зарастании вырубок пихтарников Енисейского кряжа // Почвоведение. 2015. № 8. С. 4–16.
Coxson D.S., Parkinson D. Winter respiratory activity in aspen woodland forest floor litter and soils // Soil Biol. Biochem. 1987. V. 19. P. 49–59.
Ellenberg H. Zeigerwerte der Gefasspflanzen Mitteleuropas. Gottingen: Goltze, 1974. 97 p.
Landolt E., Bäumler B., Erhardt A. Flora indicative. Ökolo-gische Zeigerwerte und biologische Kennze-ichenzur Flora der Schweiz und der Alpen. Haupt-Verlag, 2010. 376 p.
Lõhmus K., Rosenvald K., Sõber A. Elevated atmospheric humidity shapes the carbon cycle of a silver birch forest ecosystem: A FAHM study // Sci. Total Environ. 2019. V. 661. P. 441–448. https://doi.org/1.1016/j.scitotenv.2019.01.160
Middleton M., Närhi P., Arkimaa H., Hyvönen Е., Kuosmanen V., Treitz P., Sutinen R. Ordination and hyperspecrtal remote sensing approach to classify peatland biotopes along soil moisture and fertility gradients // Remote Sensing Environ. 2012. V. 124. P. 596–609.
Nikula S., Vapaavuori E., Manninen S. Urbanization-related changes in European aspen (Populus tremula L.): Leaf traits and litter decomposition // Environ. Poll. 2010. V. 158. № 6. P. 2132–2142. https://doi.org/10.1016/j.envpol.2010.02.025
Rogers P.C., Pinno B.D., Kulakowski D. A global view of aspen: Conservation science for widespread keystone systems // Global Ecology and Conservation. 2020. V. 21. https://doi.org/10.1016/j.gecco.2019.e00828
Rosenvald K., Lõhmus K., Tullus A. The initial overreaction of carbon cycle to elevated atmospheric humidity levels off over time – a FAHM study in a young birch forest // Sci. Total Environ. 2021. V. 796. https://doi.org/10.1016/j.scitotenv.2021.148917
Sekretenko O.P., Efremova T.T., Efremov S.P. Factors influencing the spatial pattern of the ash content of bog birch forest litter // Procedia Environ. Sci. 2011. V. 3. P. 99–104. https://doi.org/10.1016/j.proenv.2011.02.018
Uri V., Kukumägi M., Karoles K. Litterfall dynamics in Scots pine (Pinus sylvestris), Norway spruce (Picea abies) and birch (Betula) stands in Estonia // Forest Ecology and Management. 2022. V. 520. P. 120417–120424. https://doi.org/10.1016/j.foreco.2022.120417
Wang Q., Kwak J.-H., Chang S.X. Long-term N and S addition and changed litter chemistry do not affect trembling aspen leaf litter decomposition, elemental composition and enzyme activity in a boreal fores // Environ. Poll. 2019. V. 250. P. 143–154. https://doi.org/10.1016/j.envpol.2019.04.007
Дополнительные материалы отсутствуют.
Инструменты
Почвоведение