

Лесоведение, 2020, № 5, стр. 442-450
Оптимальный возраст оценки генетических свойств плюсовых деревьев в испытательных культурах ели европейской
А. С. Бондаренко a, *, А. В. Жигунов b, **
a Санкт-Петербургский научно-исследовательский институт лесного хозяйства
194021 Санкт-Петербург, Институтский просп., 21, Россия
b Санкт-Петербургский государственный лесотехнический университет им. С.М. Кирова
194021 Санкт-Петербург, Институтский пер., 5, Россия
* E-mail: asbond@mail.ru
** E-mail: a.zhigunov@bk.ru
Поступила в редакцию 28.12.2016
После доработки 27.02.2018
Принята к публикации 06.04.2020
Аннотация
Момент стабилизации рангового положения отдельных деревьев и полусибсовых семей плюсовых деревьев в испытательных культурах позволяет определить оптимальный возраст оценки генетических свойств древесных растений по семенному потомству. Отдельные деревья, а также полусибсовые семьи плюсовых деревьев ели европейской в насаждении характеризуются интенсивной сменой своего рангового положения в первые 10–15 лет жизни, а в последующие годы его скорость существенно снижается. Начиная со второго класса возраста (21 год и старше) наблюдается относительная стабилизация данного процесса по скорости роста. В возрасте, превышающем возраст окончательной оценки семей плюсовых деревьев в испытательных культурах (для ели европейской – 40 лет), изменения рангового положения семей также имеют место, но они гораздо менее интенсивные. Группа средних по скорости роста полусибсовых семей плюсовых деревьев характеризуется более интенсивной сменой своего рангового положения по сравнению с семьями, относящимися к крайним ранговым группам. По результатам исследований рекомендуется снижение возраста оценки генетических свойств плюсовых деревьев ели европейской в условиях Северо-Запада России до 20 лет.
При создании лесосеменных плантаций основных лесообразующих пород непроверенным по потомству материалом, генетический эффект зависит от породы, селектируемого признака и интенсивности селекции. Он составляет по разным данным от 3 до 12% для высоты дерева и от 0 до 30% – для объема ствола (Ruotsalainen, 2014). Для получения более выраженного селекционного эффекта необходимо выполнить генетическую оценку плюсовых деревьев по семенному потомству. Для оценки наследственных свойств плюсовых деревьев используются испытательные культуры, представляющие собой заложенные по специальным схемам опытные объекты, создаваемые с использованием их семенного потомства. Цель создания таких объектов – оценка преимущества в росте семенного потомства тех или иных генотипов (Основные …, 1982), которая выполняется в возрасте, соответствующем стабилизации рангового положения деревьев относительно друг друга. Именно в это время исследователи делают вывод о генетически обусловленном преимуществе в росте тех или иных семей плюсовых деревьев. По результатам исследования семьи с хорошей сохранностью растений, показавшие достоверное превышение в скорости роста над контролем, подлежат переводу в категорию элитных (Жигунов и др., 2012). В дальнейшем вегетативным потомством элитных деревьев закладываются лесосеменные плантации второго порядка. В соответствии с “Указаниями по лесному семеноводству в Российской Федерации” (2000), окончательная оценка семенных потомств плюсовых деревьев в испытательных культурах производится в возрасте потомств не менее 1/2 возраста рубки главного пользования, или возраста спелости, принятого для данного вида лесных растений в конкретной лесорастительной зоне. При этом основным вопросом в испытании генотипов лесообразующих пород является оценка уровня генетической обусловленности процессов роста и дифференциации растений в древостое по данному показателю (Маслаков, 1984; Бондаренко, Жигунов, 2007).
В частности, пристальное внимание исследователей привлекает возраст стабилизации рангового положения деревьев в насаждении, дающий ориентировочную оценку сроков испытания ценных генотипов. При этом оценки возраста, при котором происходит стабилизация рангов высот, позволяющая производить сравнительную оценку их роста и, следовательно, предсказывать преобладание в будущем по высоте тех или иных деревьев, существенно различаются. В данном контексте исследователи указывают диапазон возрастов от 5–7 до 80 лет (Картель, Манцевич, 1970; Долголиков, 1974; Орленко, 1974; Роне, 1980; Этверк, 1981; Jiang, 1987; Ворончихин, Видякин, 1989; Ефимов, 2000; Царев, 2003; Золотой, Казей, 2006; Федорков, 2009; Жигунов и др., 2012). Чрезвычайно высокий разброс оценок минимального для сравнительной характеристики генотипов возраста, очевидно, связан в первую очередь с различным подходом исследователей к допустимой точности результатов работы и формированию критериев такой оценки. Существуют различные методические подходы к моделированию процессов роста (Роне, 1980; Маслаков, 1984; Бондаренко, 2000; Демаков 2002; Xie, 2006), которые возможно использовать при изучении дифференциации деревьев в древостое.
При изучении скорости роста в высоту обычно всю совокупность деревьев исследуемой популяции разделяют на пять групп в зависимости от их высоты в определeнном возрасте (Маслаков, 1984). Недостатком такого деления деревьев является нестабильность во времени рангового положения дерева в популяции, что усложняет отнесение его к какой-либо ранговой группе. В связи с этим очевидна необходимость использовать данные о росте деревьев на всем изучаемом промежутке, а не только за какой–либо отдельный год наблюдения, что становится возможным при разделении деревьев на группы роста с помощью кластерного анализа (Бондаренко, 2000; Наследов, 2013). Следует отметить, что применение такого многомерного метода статистического анализа данных, как кластерный анализ, обусловлено исключительно удобством разделения объектов (растений) по совокупности численных характеристик (данные высот за ряд лет), не лимитировано использованием каких-либо критериев соответствия и с таким же успехом могло быть выполнено в ручном режиме самим исследователем. То есть метод кластерного анализа “К-средних” используется в качестве рабочего инструмента исследования и не включает в свой состав каких-либо статистических гипотез, на соответствие которым проверяются эмпирические данные.
При этом степень генетической обусловленности скорости роста в значительной мере определяет возможность разработки и использования методов ранней диагностики деревьев, выделенных по данному показателю, что является одним из приоритетных направлений лесовосстановления (Шутов и др., 1984; Долголиков, 1987; Беляев, 1999). В связи с этим необходима оценка возраста стабилизации рангового положения отдельных деревьев и полусибсовых семей плюсовых деревьев в целом, которая позволит определить оптимальный возраст для оценки генетических свойств древесных растений в испытательных культурах.
ОБЪЕКТЫ И МЕТОДИКА
Объектами исследования послужили испытательные культуры ели европейской, заложенные на лесных участках Гатчинского лесничества Ленинградской области в период с 1968 по 1993 г. Испытательные культуры плюсовых деревьев ели европейской используются по той причине, что для них характерны некоторые специфические особенности, делающие их идеальными объектами для выполнения такого рода исследований. В частности, они закладываются по общепринятой в России схеме для лесных культур, схема закладки участков позволяет выполнить генетический анализ, уровень генетического разнообразия семенных потомств характеризует значение данного показателя для семян с улучшенными наследственными свойствами. Перечисленное дает возможность делать выводы об особенностях роста и дифференциации производственных лесных культур на основе изучения испытательных культур.
На каждом из участков испытательных культур выполнен подеревный учет с измерением основных биометрических показателей. Точность измерения высоты растений – до 1 см на участках моложе 20-летнего возраста и до 10 см – на участках старшего возраста. Измеряется также таксационный диаметр на высоте груди (1.3 м) с точностью до 1 мм. На участке испытательных культур ели европейской в кв. 48 Орлинского участкового лесничества Гатчинского лесничества в связи с высокой густотой сплошное измерение высот деревьев выполнить затруднительно, поэтому этот показатель замерен у части деревьев числом от 2 до 13 для каждой из семей. Для расчёта средней высоты каждой из семей плюсовых деревьев использована линейная зависимость высоты от диаметра, поскольку в начальный период хода роста для деревьев ели европейской характерна линейная связь этих показателей. При этом производится индивидуальный расчет коэффициентов регрессии для каждой из полусибсовых семей, представленных на данном участке. По результатам вычисления линейной регрессии для каждой семьи получены значения коэффициентов регрессии и рассчитаны соответствующие высоты для всех исследованных плюсовых деревьев в рамках соответствующих семей. В изучаемых культурах семьи плюсовых деревьев представлены числом повторностей от 1 до 12.
Для оценки роста деревьев в испытательных культурах использовалось разбиение всей совокупности растений на ранговые группы в соответствии с их высотой. При этом деревья объединяются в возможно более удаленные друг от друга группы (кластеры) по совокупности численных характеристик. В частности, для решения приведённой задачи использовался метод кластеризации “К-средних”, при котором исследователем изначально задаётся число кластеров, на которые необходимо разделить объекты. В нашем случае объектами изучения являются деревья в испытательных культурах, кластерами – пять групп скорости роста, переменными – высоты за ряд лет. Результатом анализа является разделение деревьев на группы роста по данным ряда высот за определенный промежуток времени. По результатам выполненной группировки растений различия между группами роста в разном возрасте оцениваются при помощи дисперсионного анализа (Айвазян, 1983). Рассчитывается уровень достоверности различий между полученными группами по среднему значению высоты, а также анализируется динамика различий между полученными группами роста по высоте в возрастном градиенте. При выполнении такого анализа помимо различий между группами роста по значениям изучаемого признака рассматриваются таблицы численного распределения деревьев по группам роста, характеризующие интенсивность перехода деревьев из одних групп в другие. Доля деревьев, переходящих из одних групп в другие за определенный промежуток времени, позволяет оценить степень стабильности рангового положения деревьев разного размера в различном возрасте.
Одним из важнейших вопросов исследования являлась оценка влияния генетических факторов на стабильность проявления признака скорости роста в высоту. При этом рассматривается стабильность средних семейственных значений высоты растений в различном возрасте и доля семей, сохранивших свой ранг по высоте растений в ряду других семей. Кроме того, большой интерес представляет направление изменений рангового статуса семей. При этом внимание уделяется всем возрастным периодам развития испытательных культур ели европейской вплоть до окончания оценки их генетических свойств, что в отношении данного вида на Северо-Западе России соответствует возрасту насаждений 40 лет и старше. Для возможности выполнения сравнительного анализа скорости роста в высоту семей плюсовых деревьев в различные периоды времени использовали такой относительный показатель как редукционное число по высоте ствола, представляющее собой отношение средней высоты той или иной семьи к среднему значению высоты всего насаждения (Маслаков, 1984).
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
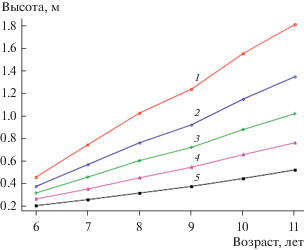
При разделении совокупности деревьев участка испытательных культур ели европейской в кв. 4 Орлинского участкового лесничества Гатчинского лесничества на пять групп роста при помощи кластерного анализа по данным ряда высот растений в возрастном периоде 6–11 лет получены хорошо обособленные группы роста (рис. 1). При этом в соответствии с результатами дисперсионного анализа различия между группами уже в возрасте растений 6 лет достоверны на уровне значимости 0.05, и эти различия между группами с возрастом увеличиваются. Так, к возрасту растений 23 г. различия между средними значениями высот растений указанных групп достигают очень высоких величин (F-критерий Фишера равен 5681). Это подтверждает предположение, что уже начиная с первых лет роста в насаждениях ели европейской наблюдается выраженная дифференциация растений по скорости прироста в высоту и с течением времени различия по высоте между деревьями-лидерами, средними деревьями и аутсайдерами только увеличивается (рис. 1).
Рис. 1.
Изменение высот растений различных групп роста (1–5) по результатам разделения совокупности растений на основе кластерного анализа “K-means” (испытательные культуры ели европейской, Гатчинское лесничество, Орлинское участковое лесничество, кв. 4).
Представление о ранговых перемещениях отдельных деревьев в древостое дают диаграммы перехода деревьев из одних групп роста в другие за определенный промежуток времени. На рисунке 2 приведен пример такой диаграммы для совокупности деревьев изучаемого участка испытательных культур ели европейской. Они отражают число деревьев по группам роста в возрасте культур 23 г. из тех, которые в соответствии с группировкой на основе кластерного анализа были отнесены по высотам растений в промежутке 6–11 лет соответственно к 1-, 3- и 5-й группам роста. В частности, из общего числа деревьев, которые по динамике хода роста в 6–11 лет по результатам кластерного анализа были отнесены к 1-й группе роста, к возрасту насаждения 23 г. в 1-2-й группах роста остались свыше 92% деревьев, а перешли в 4–5-ю группы – менее 5% деревьев (рис. 2а).
Рис. 2.
Распределение растений в возрасте 23 лет по группам роста из различных исходных групп роста, рассчитанных кластерным анализом по совокупности высот в возрасте 6–11 лет.
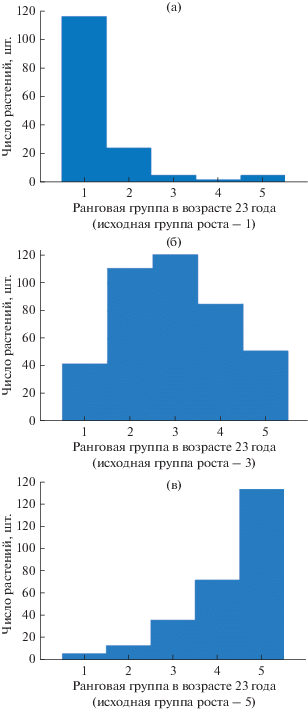
Из общего числа деревьев, которые по результатам анализа динамики хода роста в 6–11 лет были отнесены к 3-й группе роста, к возрасту насаждения 23 г. остались в 3-й группе 30% деревьев, в пределах средних 2–4-й групп роста – 78%, изменили свою группу роста на 1-ю – 10%, переместились в аутсайдеры – 12% деревьев (рис. 2б).
Если же рассматриваем исходную в возрасте 6–11 лет 5-ю группу роста (рис. 2в), то к возрасту насаждения 23 г. остались в ней осталось – 55% деревьев, сохранили положение ниже среднего (3–5-я группы) – 93%, перешли во 2-ю группу – 5%, перешли в лидирующую 1-ю группу всего 2% деревьев.
Необходимо отметить, что в соответствии с полученными данными группа средних по скорости роста полусибсовых семей плюсовых деревьев гораздо более изменчива по своему ранговому статусу, чем крайние группы роста, относящиеся к лидерам и аутсайдерам (соответственно 1- и 5-я ранговые группы).
Следует отметить, что при построении данных графических зависимостей использовались только деревья, сохранившиеся к 23 г. Очевидно, что при включении в такого рода анализ особей, которые к этому времени погибли, картина дифференциации деревьев по скорости роста была бы ещё более выраженной. Таким образом, наблюдается высокая степень стабильности рангового положения отдельных деревьев ели европейской уже после 10-летнего возраста.
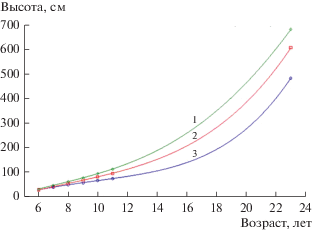
С целью оценки роли генетических факторов в процессах дифференциации растений рассмотрим динамику ранговых перемещений не по высотам отдельных деревьев, как это было сделано в приведенных выше примерах, а по уровню стабильности рангов средних значений высот по семьям плюсовых деревьев, то есть их различных генетических групп. Для этого применим рассмотренный выше способ распределения растений по группам роста при помощи кластерного анализа, но с использованием значений высот не по отдельным растениям, а средних значений высот различных полусибсовых семей плюсовых деревьев в испытательных культурах. В отличие от используемого выше разделения отдельных растений выполнено деление всего на три группы роста (лучшие, средние и аутсайдеры). Применение в анализе меньшего числа групп роста связано со сравнительно небольшой численностью объектов при кластерном анализе (целые семьи, а не отдельные деревья). Результаты такого распределения полусибсовых семей плюсовых деревьев в испытательных культурах ели европейской по трем группам роста приведены на рис. 3. Также, как и в случае с отдельными деревьями, наблюдается отчетливая дифференциация по группам роста. При этом различия между приведенными тремя группами роста в соответствии с результатами дисперсионного анализа достоверны уже начиная с 7-летнего возраста растений и последовательно увеличиваются с возрастом. Так, в возрасте растений 6 лет F-критерий равен 3.25 (значение недостоверно на уровне значимости 0.05), в возрасте 7 лет он составляет уже достоверную величину 4.63 и в дальнейшем закономерно увеличивается вплоть до различий на очень высоком уровне значимости со значением 67.01 в возрасте 23 г.
Рис. 3.
Изменение средних высот растений в семьях плюсовых деревьев ели европейской по группам роста (1–3), выделенных по результатам кластерного анализа (испытательные культуры, Гатчинское лесничество, Орлинское участковое лесничество, кв. 4).
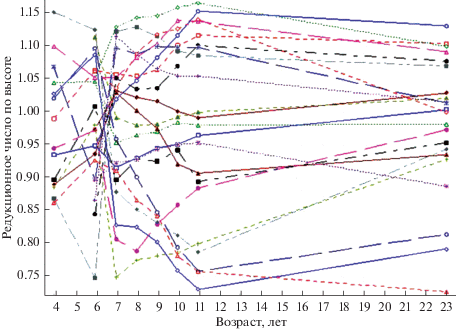
Для того, чтобы оценить стабильность рангового положения семей, рассмотрим картину динамики редукционных чисел для отдельных семей плюсовых деревьев в возрасте растений от 4 до 23 лет. В соответствии с графиком изменения редукционных чисел по высоте растений для отдельных семей плюсовых деревьев наиболее интенсивная смена их рангового положения наблюдается в возрасте до 7 лет (рис. 4). В дальнейшем ранговое положение семей на площади участка гораздо более стабильно: существенным образом меняют свои ранги только некоторые из них. Например, семья 455 в возрасте 6 лет характеризуется редукционным числом 1.10, в то время как к возрасту растений 11 лет значение данного показателя неуклонно снижается до 0.76. И всё же основная масса семей после возраста растений 7–8 лет стабилизирует свое положение в древостое по среднему значению высоты.
Рис. 4.
Динамика редукционных чисел по семьям плюсовых деревьев ели европейской (испытательные культуры, Гатчинское лесничество, Орлинское участковое лесничество, кв. 4).
Анализируя ранговые перемещения семей плюсовых деревьев на протяжении 19 лет (возраст с 4 до 23 лет) на участке испытательных культур ели европейской, расположенных в кв. 4 Орлинского участкового лесничества Гатчинского лесничества, можем говорить о том, что из одиннадцати быстрорастущих семей, имеющих в возрасте 8 лет редукционное число, превышающее 1.0 (среднее значение высоты насаждения в целом), к возрасту 23 г. (т.е. за 15 лет) сохранили свой статус 10. Таким образом, из числа семей, равных либо превышающих по высоте среднее дерево насаждения, за 15 лет роста (с 8 до 23 лет) свой лидирующий статус сохранили более 90%.
В то же время в более ранний период возрастов – с 6 до 8 лет (т.е., всего за двухлетний промежуток времени) – из 10 семей сохранили редукционное число, равное 1.0 и более только 5 семей, то есть всего лишь 50% от общего числа. Полученная картина подтверждается приведенными выше результатами анализа динамики ранговых перемещений семей плюсовых деревьев, демонстрирующими интенсивную смену рангового положения семей в возрасте 4–8 лет и гораздо большую стабильность относительного положения семей по высоте растений в последующие годы (рис. 4) – уже к 8-летнему возрасту: так, из одиннадцати семей, имеющих в возрасте 8 лет редукционное число, равное 1.0 и более, к возрасту 11 лет сохранили свое лидирующее положение 9 (81%). Если же мы рассматриваем стабильность рангового положения полусибсовых семей по высоте в возрастном промежутке от 11 до 23 лет, то за указанные 12 лет из 10 семей, имеющих в возрасте 11 лет редукционное число, равное 1.0 и более, к возрасту 23 г. сохранили свое лидирующее положение все 10 семей, то есть 100%! Таким образом, на основании приведенных данных можно заключить, что после 11 лет ранговое положение деревьев изменяется на порядок менее интенсивно, чем в первое десятилетие жизни.
Для того, чтобы оценить динамику рангового положения полусибсовых семей плюсовых деревьев воспользуемся данными о ходе роста деревьев ели европейской на одном из старейших участков испытательных культур, расположенных в кв. 48 Орлинского участкового лесничества Гатчинского лесничества Ленинградской области. По результатам аналогичного вышеприведенному анализа рангового положения семей плюсовых деревьев в возрастном градиенте из семи лучших семей, имеющих в возрасте 18 лет редукционное число равное 1.0 и более, к возрасту 49 лет (то есть в течение 1–2-го классов возраста за 31 г. наблюдений) сохранили свое лидирующее положение 5 из них, т.е. 71%. Если же мы используем более поздний промежуток времени, соответствующий первой половине 2-го класса возраста, то в возрасте от 44 до 49 лет из числа лучших, изменила свое лидирующее положение (редукционное число, равно 1.0 и более) только одна семья плюсового дерева 36–54.
Таким образом, в возрасте окончательной оценки наследственных свойств плюсовых деревьев (в соответствии с действующими нормативными документами для ели европейской это 40 лет) смена рангового положения семей плюсовых деревьев все еще имеет место, хотя и выражена в гораздо меньшей степени по сравнению с периодом возрастов до середины 2-го класса.
Основываясь на выполненных исследованиях можно говорить, что в конце 2-го класса возраста в насаждениях ели европейской в условиях Северо-Запада России с достаточно высокой долей вероятности становится возможной оценка наследственных свойств растений по скорости роста в испытательных культурах. Интенсивная смена рангового положения отдельных деревьев и полусибсовых семей плюсовых деревьев наблюдается в раннем возрасте в течение первой половины первого класса возраста. Начиная со второго класса возраста (21 год и старше) наблюдается относительная стабилизация рангового положения семей плюсовых деревьев по скорости роста. В связи с этим полученные результаты могут быть использованы в качестве одного из обоснований для снижения возраста оценки генетических свойств плюсовых деревьев ели европейской в условиях Северо-Запада России. Это позволит сократить сроки испытания генотипов основных лесообразующих пород, смягчить негативные последствия старения вегетативного материала при закладке лесосеменных плантаций, обеспечить необходимую преемственность работ в лесной селекции и повысить экономическую эффективность селекционных работ за счет интенсификации использования селекционного материала при производстве лесных культур.
Выводы. 1. Отдельные деревья ели европейской в насаждении характеризуются интенсивной сменой своего рангового положения в первые 10–15 лет жизни.
2. Наиболее интенсивная смена рангового положения у полусибсовых семей плюсовых деревьев ели европейской наблюдается в возрасте до 10 лет. В дальнейшем интенсивность смены рангового положения семей плюсовых деревьев существенно снижается. Тем не менее некоторые изменения рангового положения семей плюсовых деревьев наблюдаются и в возрасте, превышающем возраст окончательной оценки семей плюсовых деревьев в испытательных культурах (для ели европейской – 40 лет).
3. Группа средних по скорости роста полусибсовых семей плюсовых деревьев характеризуется более интенсивной сменой своего рангового положения в древостое по сравнению с семьями, относящимися к крайним ранговым группам (то есть лидерам и аутсайдерам, соответственно 1- и 5-я ранговые группы).
4. Начиная со второго класса возраста (21 год и старше) наблюдается относительная стабилизация рангового положения семей плюсовых деревьев ели европейской по скорости роста.
5. Рекомендуется снижение возраста оценки генетических свойств плюсовых деревьев ели европейской в условиях Северо-Запада России до 20 лет для обеспечения снижения общего срока испытания генотипов и закладки лесосеменных плантаций второго порядка.
Список литературы
Айвазян С.А., Енюков И.С., Мешалкин Л.Д. Прикладная статистика: Исследование зависимостей. М.: Финансы и статистика, 1983. 487 с.
Беляев В.В. Эффективность отбора посадочного материала хвойных пород по прямому и косвенным признакам при создании лесных культур // Лесной журн. 1999. № 6. С. 41–48.
Бондаренко А. С. Применение кластерного анализа для оценки роста древесных пород // Восстановление лесов, ресурсо- и энергосберегающие технологии лесного комплекса: Мат. межвузовс. науч.-практ. конф., посвященной 70-летию Воронежской лесотех. академии. Воронеж, 27–29 сентября 2000 г. Воронеж, 2000. С. 40–42.
Бондаренко А.С., Жигунов А.В. Генетическая обусловленность скорости роста ели европейской в культуре // Лесоведение. 2007. № 1. С. 42–48.
Ворончихин Л.И., Видякин А.И., Овечкин С.М. Пути повышения эффективности работ по селекции сосны и ели в Кировской области // Селекция ценных форм древесных пород и их использование для создания целевых насаждений: Сб. науч. тр. Воронеж: Центральный НИИ лесной генетики и селекции, 1989. С. 79–83.
Демаков Ю.П. Изменчивость и классификация форм кривых роста деревьев в онтогенезе // Лесной журн. 2002. № 4. С. 34–41.
Долголиков В.И. О ранней диагностике быстрого роста в высоту у сосны и ели по прямому признаку // Состояние и перспективы развития лесной генетики, селекции, семеноводства и интродукции. Методы селекции древесных пород: Сб. тез. докл. совещ. Рига: Зинатне, 1974. С. 45–48.
Долголиков В.И. Отбор быстрорастущих саженцев ели для плантационного лесовыращивания: Методич. указания. Л.: ЛенНИИ лесн. хоз-ва, 1987. 20 с.
Ефимов Ю.П. Итоги многолетнего испытания материнских деревьев сосны обыкновенной по семенному потомству // Генетическая оценка исходного материала в лесной селекции. Воронеж: НИИ лесной генетики и селекции, 2000. С. 33–44.
Жигунов А.В., Бондаренко А.С., Николаева М.А. Первые результаты отбора элитных деревьев ели европейской в Ленинградской области // Лесной журн. 2012. № 3. С. 43–50.
Золотой А.В., Казей Т.П., Яблоков С.А. Селекционная оценка испытательных культур ели европейской в ОЛХ “Русский лес” // Лесной вестник. 2006. № 5. С. 135–138.
Картель Н.А., Манцевич Е.Д. Генетика в лесоводстве. Минск: Наука и техника, 1970. 165 с.
Маслаков Е.А. Формирование сосновых молодняков. М.: Лесная пром-сть, 1984. 168 с.
Наследов А. IBM SPSS Statistics 20 и AMOS: профессиональный статистический анализ данных. СПб.: Питер, 2013. 416 с.
Орленко Е.Г. Ранняя диагностика при проверке генетических свойств плюсовых деревьев // Состояние и перспективы развития лесной генетики, селекции, семеноводства и интродукции: Сб. тез. докл. совещ. Рига: Зинатне, 1974. С. 83–84.
Основные положения методики закладки испытательных культур плюсовых деревьев основных лесообразующих пород. Воронеж: Центральный НИИ лесной генетики и селекции, 1982. 18 с.
Роне В.М. Генетический анализ лесных популяций. М.: Наука, 1980. 160 с.
Указания по лесному семеноводству в Российской Федерации. Утв. Рослесхозом 11.01.2000 г. М: ВНИИЦлесресурс, 2000. 198 с.
Федорков, А.Л., Туркин А.А. Возраст оценки качества потомств в испытательных культурах сосны // Лесоведение. 2009. № 2. С. 69–71.
Царев А.П., Погиба С.П., Тренин В.В. Селекция и репродукция лесных древесных пород. М.: Логос, 2003. 520 с.
Шутов И.В., Маслаков Е. Л., Маркова И.А. Лесные плантации (ускоренное выращивание ели и сосны). М.: Лесная пром-сть, 1984. 245 с.
Этверк И.Э. Результаты испытания потомств елей одного насаждения // Разработка основ систем селекции древесных пород: Тез. докл. совещ. Рига, 22–25 сентября 1981 г. Рига: Зинатне, 1981. Ч. 1. С. 122–125.
Jiang I. B-J. Early testing in forest tree breeding: a review // Forest Tree Improvement. 1987. № 20. P. 45–78.
Ruotsalainen S. Increased forest production through forest tree breeding // Scandinavian J. Forest Research. 2014. V. 29. № 4. P. 333–344
Xie C.-Y., Yanchuk A.D., Fu. Y.-B. Accuracy of ranking Individuals in field tests of different designs: acomputer simulation /// Silvae Genetica. 2006. V. 55. № 2. P. 70–77.
Дополнительные материалы отсутствуют.