БИОФИЗИКА, 2023, том 68, № 5, с. 1046-1056
БИОФИЗИКА CЛОЖНЫX CИCТЕМ
УДК 577.3
СЛОЖНАЯ СИСТЕМА МОЗГА МЫШИ - СЕЛЕКЦИЯ НА УСПЕШНОСТЬ
РЕШЕНИЯ КОГНИТИВНОГО ТЕСТА
© 2023 г. И.И. Полетаева*, #, О.В. Перепелкина*, З.А. Зорина*
*Биологический факультет Московского государственного университета имени М.В. Ломоносова,
Ленинские горы, 1/12, Москва, 119234, Россия
#E-mail: ingapoletaeva@mail.ru
Поступила в редакцию 26.06.2023 г.
После доработки 03.07.2023 г.
Принята к публикации 22.09.2023 г.
Представлены экспериментальные данные по положительным результатам селекционного экспе-
римента у мышей. Отбор вели на успешное решение когнитивного теста (требующего от животного
оперирования правилом «неисчезаемости» предметов) и на не-решение этого теста. Полученные
данные показали, что у мышей линии, селектированной на успешное решение теста, были досто-
верно лучше выражены краткосрочная рабочая память, а также показатели тестов на новизну и на
внимание. Это позволяет предполагать, что межлинейные различия касаются не только способно-
сти к решению данного теста, а связаны с более четко выраженными «исполнительными функци-
ями».
Ключевые слова: когнитивные способности животных, элементарная логическая задача, генетика по-
ведения, селекция, лабораторные мыши.
DOI: 10.31857/S0006302923050265, EDN: NAWPTG:
Работа посвящается памяти Г.В. Гурского
Мозг - сложная система, причем некоторые
ях в публикациях последних лет - это, преимуще-
полагают, что это «самая сложная» система орга-
ственно, сообщения об отличии групп животных
низма. Особенно высока сложность мозга челове-
«нормального контроля» («дикого типа») от пока-
ка. В связи с этим большое значение имеют ис-
зателей тестов животных либо с измененным ге-
следования когнитивных способностей на лабо-
номом, либо с особенностями физиологического
раторных животных, поскольку эти объекты и
состояния вследствие разрушений мозга или дей-
проще по строению, и позволяют оценивать
ствия фармакологических агентов.
большое число нейрофизиологических призна-
Изменчивость признаков поведения, которую
ков, связанных с когнитивными способностями.
обнаруживают животные практически во всех ис-
Вторая половина XX века и начало XXI века озна-
пользуемых тестах (и, как мы знаем, в «обычном»
менованы накоплением данных (часто уникаль-
поведении), имеет генетическую и средовую со-
ных) о возможностях центральной нервной си-
ставляющие, а также составляющую, связанную с
стемы (ЦНС) и когнитивных способностях жи-
генотип-средовыми взаимодействиями. Выпол-
вотных многих видов
- от дрозофилы
[1],
нение животным тех или иных актов поведения
муравьев [2], грызунов [3] до человекообразных
(как в лабораторных тестах, так и в природе) ос-
обезьян [4] (возможности мозга человека здесь не
новано их на врожденной основе и их сложной
рассматриваются).
модуляции при накоплении опыта. Взаимодей-
ствие генотипических и средовых влияний - это
Анализ современных исследований способно-
важный элемент понимания механизмов поведе-
стей животных решать сложные «интеллектуаль-
ния, а для этого, однако, важно учитывать это со-
ные» задачи (например, сложные задачи на про-
странственную ориентацию и др.) показывает,
отношение («врожденного» и «приобретенного»)
что основное внимание в настоящее время при-
в репертуаре поведения конкретной группы жи-
вотных (см. напр., [7]).
влечено к созданию моделей (чаще всего, генети-
ческих моделей) заболеваний мозга человека [5,
Важным моментом в оценке относительной
6] и др. В то же время биологические основы
роли «врожденного» и «приобретенного» в адап-
функционирования мозга в нормальных услови-
тивных поведенческих реакциях является следу-
1046
СЛОЖНАЯ СИСТЕМА МОЗГА МЫШИ
1047
ющее. Основные элементы врожденных (ин-
tor. В устойчивых к заболеванию колониях рабо-
стинктивных) движений (спонтанных или прояв-
чие пчелы, обнаружив (видимо, по запаху) ячей-
ляющихся в ответ на определенные сигналы) -
ки с пораженными болезнью личинками,
это так называемые «фиксированные комплексы
вскрывают их и выбрасывают больных личинок
действий» (ФКД) - видоспецифические последо-
из улья. Классический генетический экспери-
вательности движений.
мент позволил выявить эффект двух генов, один
из которых «отвечает» за поведение «вскрытия
Эти ФКД проявляются у всех особей вида в
ячейки» (специфические движения, ФКД), а вто-
«готовом» виде, без научения, они имеют специ-
рой - за удаление зараженных личинок из вскры-
фическую, выполняемую «стереотипно» форму,
тых ячеек (другие, иные по «рисунку» ФКД). Од-
практически не видоизменяясь в течение жизни
нако впоследствии выявилось, что в колониях,
(например, движения умывания или вертикаль-
подверженных инфекции, рабочие пчелы также
ные стойки грызунов). Однако консервативность
обнаруживают ФКД этих двух типов. Иначе гово-
и стереотипность ФКД находятся в видимом про-
ря, они выполняют два типа движений «гигиени-
тиворечии с принципом пластичности и измен-
ческого» поведения, но эти движения (в силу ка-
чивости поведения, которому есть огромное чис-
ких-то причин) проявляются у них очень редко и
ло подтверждений (как в лабораторных исследо-
поэтому оказываются неэффективными в плане
ваниях, так и в наблюдениях натуралистов).
устранения инфекции. Иными словами, у них
Наиболее сложным по своему механизму являет-
высок порог инициации этих движений. У пчел
ся поведение животного при научении, а также
колоний с гигиеническим поведением эти движе-
при решении элементарных логических задач,
ния инициируются легче, т.е. у них генетически
которое складывается из врожденных реакций,
детерминирован более низкий порог инициации.
выполнение которых «обусловлено» текущей сре-
Еще один пример того, как генотип может моду-
довой ситуацией.
лировать проявление поведения именно в связи с
Однако существует и путь выхода из такого ка-
уровнем возбудимости ЦНС, дали эксперименты
жущегося противоречия - изменчивыми оказы-
по наследованию пассивно-оборонительного по-
ваются не сами, иногда сложные, движения (т.е.
ведения у собак [10].
их конкретная форма), а пороги их инициации. А
Таким образом, инстинктивное поведение и
эти пороги, в свою очередь, могут испытывать
условия его экспрессии зависят от генетически
модулирующее влияние ряда факторов (темпера-
детерминированных особенностей мозговых
тура, сезон, мотивационный и гормональный
структур, обеспечивающих активацию соответ-
фон, общая возбудимость, уровень сенсорной
ствующих нервных цепей (и, по-видимому, от
стимуляции и т. д.), обнаруживая значимый уро-
числа нейронов в ряде структур мозга [11], а также
вень как межпопуляционной, так и индивидуаль-
от гормонального фона и благоприятных или, на-
ной изменчивости. К таким факторам относится
оборот, вредных для животного внешних усло-
и генотип.
вий. Основные свойства сенсомоторных меха-
На важное значение частоты проявления ФКД
низмов поведения и реактивности ЦНС на сти-
(стереотипных по паттерну) первым еще в 1967 г.
мулы предопределены в развивающейся нервной
обратил внимание выдающийся английский это-
системе, они в разной степени зависят от контак-
лог А. Меннинг, анализируя генетические разли-
тов между нейронами, а в своем фенотипическом
чия в половом поведении дрозофилы [8]. По его
проявлении обнаруживают существенных «врож-
мнению, генетическая изменчивость поведения в
денный» компонент.
большой степени определяется изменчивостью
В то же время на другом конце виртуальной
порогов провокации и, соответственно, измене-
шкалы («врожденное-приобретенное», см. работу
ниями частоты выполнения важных для адапта-
[12]) находятся феномены поведения, почти пол-
ции животного ФКД, это может также лежать в
ностью зависящие (и в формировании, и в ини-
основе микроэволюционных изменений поведе-
циации) от индивидуального опыта животного,
ния. Иными словами, эти «единицы поведения» в
хотя в своем проявлении также зависящие от
силу разных причин могут проявляться чаще или
врожденных стереотипных действий, а также от
реже (т.е. провоцироваться «легче» или «труд-
уровня развития ЦНС данного вида. Это, в част-
нее»).
ности, пластические изменения поведения при
Анализ наблюдений за поведением насеко-
процессах обучения разной сложности. Они так-
мых, грызунов и других представителей животно-
же связаны со структурными особенностями
го царства дает много материала, подтверждаю-
ЦНС, детерминированными генотипом, но связь
щего правильность высказанной Меннингом ги-
имеет очень сложный характер, поскольку в этих
потезы. Например, в знаменитой работе [9] о
случаях имеет место сложное взаимодействие
генетической основе гигиенического поведения
врожденных компонентов поведения с реакцией
пчел было показано, что колонии пчел различа-
мозга на внешние воздействия (осуществляющe-
ются по устойчивости к инфекции Varroa destruc-
еся во многих случаях на основе эпигенетических
БИОФИЗИКА том 68
№ 5
2023
1048
ПОЛЕТАЕВА и др.
механизмов). Примером могут быть различия
показано, что при использовании «батареи те-
между двумя линиями крыс, селектированными в
стов», требующих от животного научения, иногда
начале 1920 г. Р. Трайоном, различия между кото-
весьма сложного, у них можно обнаружить значи-
рыми (по данным более поздних исследований)
мую корреляцию успешности их выполнения.
имеют иногда противоречивый характер и связа-
Такой комплекс способностей также получил на-
ны также с особенностями гормонального фона
звание g-фактор (работы [17-21] и др.). Следую-
крыс этих линий. Усложнение поведения (в эво-
щим шагом в интерпретации этих данных было
люционном ряду) в этом случае идет по пути
предположение о существовании у животного
усложнения потенций, т.е. возможностей носите-
(как и у человека) совокупности способностей,
лей данной ЦНС проявить поведение, базирую-
которые были названы «исполнительные функ-
щееся на решении, например, новых задач.
ции» (executive functions). В их число входят «ко-
А. Меннинг пишет, что в нормальных условиях
гнитивная пластичность», рабочая память, спо-
ни у обезьяны, ни у крысы нет поведения, связан-
собность к предвидению, внимание, тормозный
ного с решением сложной задачи инструменталь-
контроль и др. Совокупная экспрессия высоких
ного научения, т.е. выбору определенного сочета-
показателей этих
«способностей» определяет
ния стимулов, подкрепляемого пищей. Однако
успешность целенаправленного поведения [22,
ЦНС обезьян обладает потенциями к решению
23]. При этом исследования нейрофизиологии
подобной задачи, а ЦНС крысы обладает этим в
мозга на крысах показали, например, участие в
значительно меньшей степени (см. [8]). Наличие
реализации таких способностей структур перед-
или отсутствие таких «потенций» - это свиде-
него мозга [24].
тельство способности животных данного вида к
Предметом настоящего исследования был
решению «когнитивных» задач без опыта их ре-
анализ результатов селекции лабораторных мы-
шения в прошлом.
шей на способность к решению когнитивного те-
Исследование мозговых механизмов решения
ста на неисчезаемость (по Ж. Пиаже: объект, ра-
таких задач животными разного уровня развития
нее воспринимавшийся, продолжает существо-
(т.е. задач, с которыми животное сталкивается
вать, даже если его в данный момент не видно).
впервые и, следовательно, не имеет опыта их ре-
Для решения такой элементарной логической за-
шения) - это и актуальная задача нейробиоло-
дачи животному необходимо и понимание логи-
гии, и важный в практическом отношении вопрос
ки конкретной ситуации, и внимание, и рабочая
биологических основ когнитивного поведения
память [25].
человека [13]. В этом плане важны данные по ге-
В работе представлены данные по результатам
нетическим различиям в проявлении когнитив-
отбора лабораторных мышей на высокие и низ-
ных способностей животных. Следует отметить,
кие показатели решения когнитивного теста (ос-
что в современной нейробиологической литера-
нованного на врожденной реакции мыши избе-
туре под термином «когнитивный», как правило,
гать яркое освещение). Исходной популяцией
понимается проявление пластичности поведе-
для отбора были мыши линии ЭКС (F20), селек-
ния, т.е. его изменчивости в связи с формирова-
тированной ранее [26] на успешность решения
нием индивидуального опыта. Однако в более уз-
задачи на экстраполяцию направления движения
ком понимании данного термина - это способ-
стимула, скрывшегося из поля зрения. Отметим,
ность особи решить впервые встреченную задачу
что селекционный эксперимент на способность к
на основе способности к улавливанию законо-
экстраполяции дал неустойчивые результаты, и
мерностей, связывающих предметы и явления в
можно считать, что успешной селекции на реше-
конкретной ситуации, которую Л.В. Крушин-
ние теста на экстраполяцию не произошло. Это-
ский [14] назвал «элементарной рассудочной дея-
му могут быть разные причины, анализ которых
тельностью». На примере способности животных
дан в разделе «Обсуждение». В настоящем экспе-
ряда видов к экстраполяции направления движе-
рименте признаками для отбора были показатели
ния стимула, скрывшегося из поля зрения, Кру-
успешности (линия «плюс») или неудачи (линия
шинский и его коллеги показали важность струк-
«минус») выполнения мышами теста на поиск
турной сложности ЦНС животных разных видов,
входа в укрытие (ПВУ) в его наиболее «трудных»
а также (для крыс, мышей и серебристо-черных
предъявлениях (см. ниже). Данный тест (в англо-
лисиц) были продемонстрированы генотипиче-
язычной литературе - puzzle-box test [20]) осно-
ские различия в экспрессии этого признака (см.
ван на стремлении животного спрятаться в тем-
работы [14, 15]).
ноте из ярко освещенной части эксперименталь-
Очевидно, что накопление знаний о когнитив-
ной камеры, когда путь в темноту закрыт
ных способностях животных вызвало потреб-
препятствием, которое необходимо преодолеть.
ность к их обобщению. По аналогии с представ-
Этот тест является «когнитивным» тестом per se,
лением, выдвинутым психологом К. Сперманом
поскольку для его выполнения не требуется пред-
[16] о факторе общего интеллекта (g-factor), в
варительного научения, а его решение - это ре-
большом числе исследований на грызунах было
шение элементарной логической задачи. Его
БИОФИЗИКА том 68
№ 5
2023
СЛОЖНАЯ СИСТЕМА МОЗГА МЫШИ
1049
Мышей содержали в пластиковых клетках раз-
мером 42 × 21.5 × 15 см по 6-8 особей в каждой с
естественной сменой освещенности и температу-
рой в пределах 19-22°С. Они получали стандарт-
ный корм (ООО «Лабораторкорм», Москва) и пи-
тьевую воду ad lib.
Тест на поиск входа в укрытие. Начальный ва-
риант этого теста был впервые предложен как
часть батареи тестов для оценки у мышей общего
фактора g [21, 22]. В нашем варианте теста число
предъявлений разных его этапов было укорочено
до четырех (сложность которых возрастала от
первого к третьему), а тестирование можно было
провести в течение одного опытного дня (в отли-
чие от трех дней в изначальном варианте М. Гол-
суорси и др.). Экспериментальная пластиковая
камера состоит из двух отделений - ярко осве-
Рис. 1. Схема экспериментальной установки теста на
щенного, размером 30 × 28 × 27.7 см, и темного,
поиск входа в укрытие: 1 - освещенная часть камеры,
размером 14 × 28 × 27.5 см (рис. 1, 2).
2 - темная часть камеры, 3 - углубленный в пол лаз.
Мышь, помещенная в ярко освещенную часть
экспериментальной камеры, стремится убежать и
описание представлено в разделе «Материалы и
спрятаться в ее темной части, куда ведет углуб-
методы». В добавление к оценкам способности к
ленный в пол лаз (глубина - 1.5 см, ширина -
решению этого теста на ПВУ мы оценили прояв-
4.5 см, длина - 11.5 см). В первой пробе теста лаз
ления краткосрочной рабочей памяти и внима-
открыт, во второй его маскируют чистой струж-
ния к новым предметам у этих животных.
кой, насыпая ее вровень с уровнем пола, а в тре-
тьей и четвертой пробах (которые наиболее труд-
ны мышам) лаз прикрывают и блокируют легкой
МАТЕРИАЛЫ И МЕТОДЫ
пробкой из пластика и картона, которую живот-
ное может отодвинуть или вынуть зубами из
Экспериментальные животные. Как упомина-
углубленного лаза. На решение первой и второй
лось выше, исходной популяцией для настоящего
проб животному дается по 180 с, на решение тре-
селекционного эксперимента были мыши 20-го
тьей и четвертой проб - по 240 с. После перехода
поколения селекции мышей (линия ЭКС) на
животного в темную часть камеры, его оставляют
успешность решения задачи на экстраполяцию.
в ней на 15-20 с, после чего отсаживают в отдель-
Поведение мышей линии ЭКС сравнивали с та-
ную клетку и через 45-60 с предъявляют следую-
ковым мышей неселектированной генетически
щую пробу. Регистрируется латентный период
гетерогенной популяции (КоЭКС). На основе
перехода мыши в темноту, а также ряд показате-
F20 линии ЭКС был начат новый селекционный
лей поведения, предшествовавших этому - это
эксперимент (получено 7 поколений) с отбором
латентный период первого подхода мыши к лазу,
на успешность решения теста ПВУ (линия
число подходов до решения теста, число верти-
«плюс») и на «не-решение» этого теста (линия
кальных стоек, эпизодов груминга и замирания, а
«минус»). Суммарно было протестировано более
также число болюсов дефекации. В случаях с ла-
500 мышей (самцов и самок).
зом, закрытым пробкой, при отсутствии решения
Рис. 2. Тест ПВУ: 1 - лаз открыт; 2 - лаз замаскирован стружкой, мышь ее раскапывает; 3 - мышь «в раздумье», увидев
«пробку», скрывающую лаз.
БИОФИЗИКА том 68
№ 5
2023
1050
ПОЛЕТАЕВА и др.
Рис. 3. Тест на внимание: (а) - вид камеры с подвешенным на уровне глаз мыши предметом (муляж куриного яйца); (б) -
набор использованных предметов, на которых мышь могла фиксировать внимание.
теста отмечали также наличие или отсутствие у
Регистрировали (вручную) время пребывания
мыши движений «манипулирования» пробкой,
мыши на открытой части лабиринта (открытые
т.е. попыток (безуспешных) проникнуть в темно-
рукава и центр), число выходов из темных рука-
ту - захватов пробки зубами и ее приподнимания.
вов на свет, число стоек в темных рукавах, а также
Критерии отбора. Критериями отбора для суб-
число переходов из одного темного рукава в дру-
гой.
линии «плюс» было успешное решение теста в его
наиболее «трудном» варианте (обеих проб теста с
Тест на внимание. Данный тест проводили в
«пробкой»), а для сублинии «минус» - отсутствие
круглой камере из черного пластика, диаметром
такого решения. Поскольку практически все мы-
36 см и высотой 22 см (рис. 3а). По внутренней
ши решали первую и вторую пробы теста на ПВУ
стенке камеры по ее периметру перемещали под-
и выполняли реакцию достаточно быстро, крите-
вешенный и закрепленный по краю камеры один
рием для селекции на успешное решение теста
из пяти предметов, располагавшийся на уровне
(линия «плюс») было решение третьего и четвер-
головы животного (рис. 3б). Перемещение пред-
того предъявлений теста в течение времени не
мета начинали в момент, когда голова мыши была
дольше, чем 60-90 с. В качестве родителей для
обращена к предмету (в среднем через 10-15 с по-
следующего поколения линии «минус» отбирали
сле помещения животного в установку). Каждый
животных, которые не решили этот тест при тре-
предмет демонстрировали только в течение одно-
тьем и четвертом его предъявлениях. Мышей для
го «круга» его движения, что занимало в среднем
получения потомства (один самец и одна-две
40 с. Фиксировали (вручную) время первого под-
самки) содержали в более маленьких клетках
хода к предмету (латентный период), число таких
(30 × 10.5 × 9 см), беременных самок отсаживали
подходов (приближений мыши к стенке под
в отдельные клетки такого же размера, в которых
предметом), число стоек, эпизодов груминга и за-
их содержали с потомством до прекращения ма-
мираний, а также число болюсов дефекации. В
теринского вскармливания (30-е-34-е сутки жиз-
качестве предметов, которые должны были при-
ни). После этого мышатам наносили ушные мет-
влекать внимание животных (рис. 3б) были: 1 -
ки и помещали по 6-9 особей (самцов и самок от-
светлоокрашенный муляж куриного яйца, 2 - бе-
дельно) в более крупные клетки.
лая пластиковая лопатка (диаметром 2 см), 3 - бе-
Тестирование поведения мышей начинали в
лый пластиковый диск (диаметром 2.5 см), 4 -
возрасте 2.5-3.0 мес. Каждый из использованных
темно-окрашенный (цветной) диск (диаметром
тестов проводил всегда один и тот же экспери-
3 см) и 5 - пластиковая игрушка неправильной
ментатор. Ранее было показано, что проведение
формы (2 × 2.5 см). Порядок предъявления пред-
тестирования разными лицами (в том числе и раз-
метов разным животным был случайным, а при
ного пола) влияет на показатели тестов у живот-
обработке выделяли реакции животных каждой
ных одного и того же генотипа [27, 28].
группы на предмет, показанный первым, и пред-
мет, показанный последним.
Тест приподнятого крестообразного лабиринта.
Установка располагалась на высоте 29 см над по-
Статистическая обработка. Статистическую
верхностью стола, длина открытых и закрытых
оценку достоверности различий между группами
рукавов - 21 см, ширина открытых рукавов -
животных для величин латентных периодов ре-
5 см, закрытых - 5.5 см, высота стенок закрытых
шения тестов проводили с помощью одно- и
рукавов - 18 см. Мышь помещали на 3 мин в
двухфакторного анализа ANOVA (c последующей
центр приподнятого крестообразного лабиринта.
оценкой post hoc по методу LSD Фишера). Досто-
БИОФИЗИКА том 68
№ 5
2023
СЛОЖНАЯ СИСТЕМА МОЗГА МЫШИ
1051
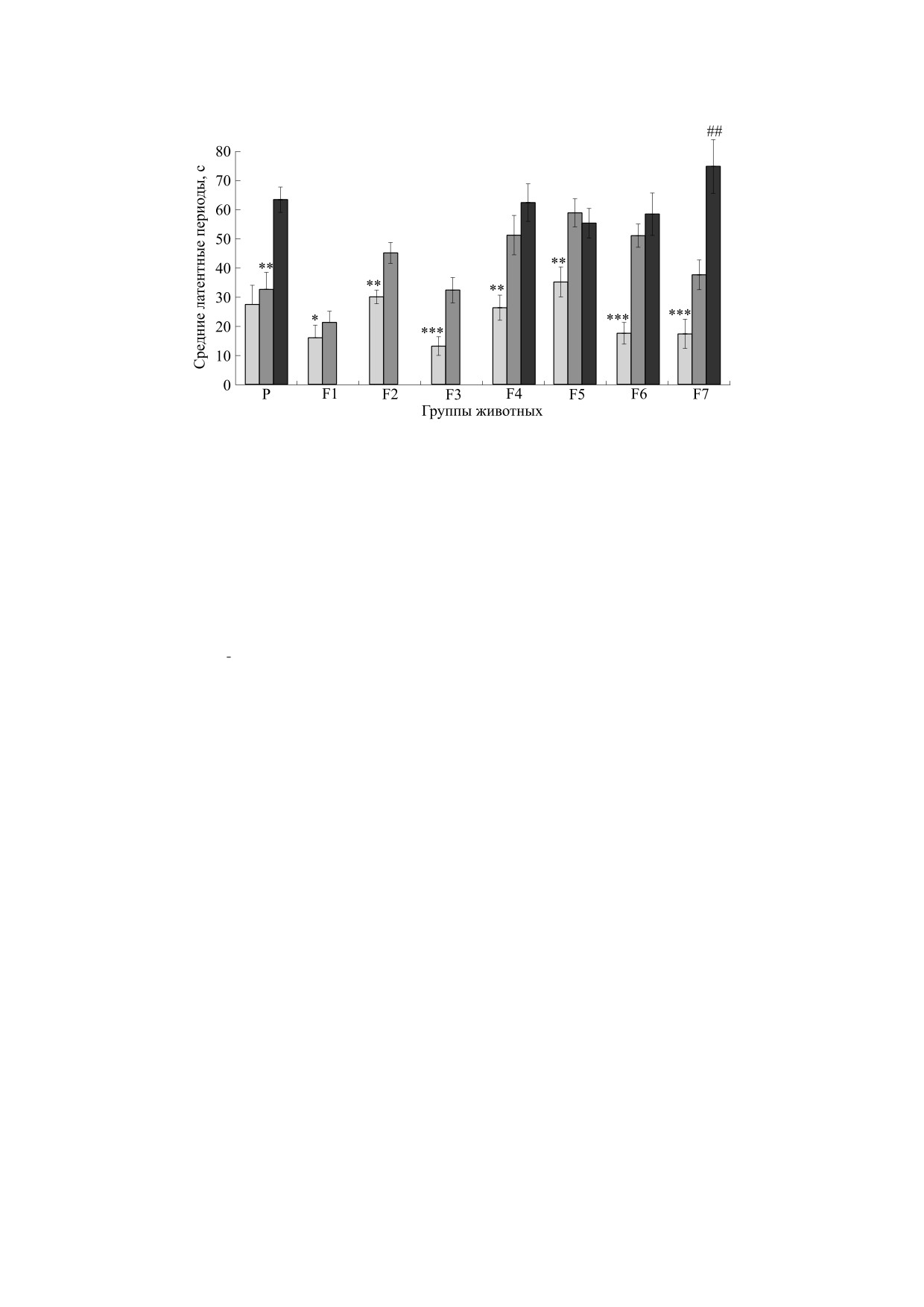
Рис. 4. Средние латентные периоды перехода в темное отделение камеры в тесте на поиск входа в укрытие мышами
разных групп, когда лаз замаскирован стружкой. По оси ординат - средние латентные периоды, с; по оси абсцисс -
группы животных: Р - мыши «родительских» пар, выбранных для последующей селекции линий «плюс» и «минус»; F1-
F7 - мыши последовательных поколений селекции и контрольной группы КоЭКС (неселектируемой генетически
гетерогенной популяции). Светло-серые столбцы - мыши линии «плюс», темно-серые столбцы - линии «минус»,
черные столбцы - мыши популяции КоЭКС; *, **, *** - достоверное отличие от показателей линии минус и от
контрольной группы, #, ## - достоверные различия между группой линии «минус» и КоЭКС при р < 0.01 (post hoc тест
LSD Фишера, однофакторный анализ ANOVA).
верность различий между долями мышей каждой
темное отделение камеры при маскировке лаза
группы, решивших и не решивших тест ПВУ,
стружкой более медленно, чем мыши «плюс».
определяли с помощью метода φ Фишера для аль-
Таким образом. выполнение теста ПВУ в пер-
тернативных долей.
вых двух пробах (лаз открыт и замаскирован
стружкой) было успешным у мышей всех групп и
достаточно быстрым, тогда как различия между
РЕЗУЛЬТАТЫ
«плюс»- и «минус»-группами мышей этих поко-
Решение теста на поиск входа в укрытие. При-
лений четко обнаруживались в предъявлениях те-
водятся результаты решения теста ПВУ, предъяв-
ста. В предъявлениях теста, когда лаз был закрыт
ленного мышам генетических групп. Это были
пробкой, выявились различия в успешности ре-
мыши последовательных поколений селекции на
шения теста (рис. 5), что может быть ответом на
решение и не решение когнитивного теста, а так-
отбор в этих поколениях. Решение теста ПВУ мы-
же (в F6 и F7) мыши контрольной неселектиро-
шами КоЭКС в третьей и четвертой пробах теста,
ванной популяции КоЭКС (n = 34) [26, 27].
когда «лаз» в темное отделение камеры закрыт
пробкой, было значительно менее успешным,
На рис. 4 представлены данные по средним ве-
чем у мышей обеих линий селекции.
личинам латентного периода решения теста ПВУ
при его первых двух предъявлениях (когда лаз от-
Наблюдения за поведением мышей показали,
крыт и лаз замаскирован стружкой) в поколениях
что в случаях, когда лаз был замаскирован проб-
F1-F7, а также мышей родительского поколения
кой, а решения теста не произошло, в большин-
линии ЭКС F20. Практически мыши всех тести-
стве случаев мыши обеих линий стараются заце-
рованных групп успешно, но с разными результа-
пить или толкнуть пробку, т.е. обнаруживают
тами по длительности латентного периода, пере-
«манипулирование» этим объектом, это наблюда-
ходили в темное отделение камеры. Обращает на
лось и у мышей F20 (исходной популяции для се-
себя внимание довольно быстрое решение теста
лекционного эксперимента). Это поведение
при лазе, замаскированном стружкой. По нашим
можно рассматривать как свидетельство того, что
наблюдениям, наличие стружки, закрывающей
мышь пытается пройти через лаз, но не может
лаз, вызывает быструю «позитивную» реакцию ее
преодолеть это препятствие, т.е. обнаруживает
раскапывания практически у всех животных (по-
«понимание» неисчезаемости искомого объекта.
скольку это, видимо, относится к репертуару
Мышей, которые не «манипулировали» пробкой,
врожденных стереотипов поведения), однако
было очень мало. Например, в F3 в сублинии
практически во всех случаях мыши, селектируе-
«плюс» было одно такое животное (из 83), в суб-
мые на «не решение» теста ПВУ, переходили в
линии «минус» - два животных (из 73). В то же
БИОФИЗИКА том 68
№ 5
2023
1052
ПОЛЕТАЕВА и др.
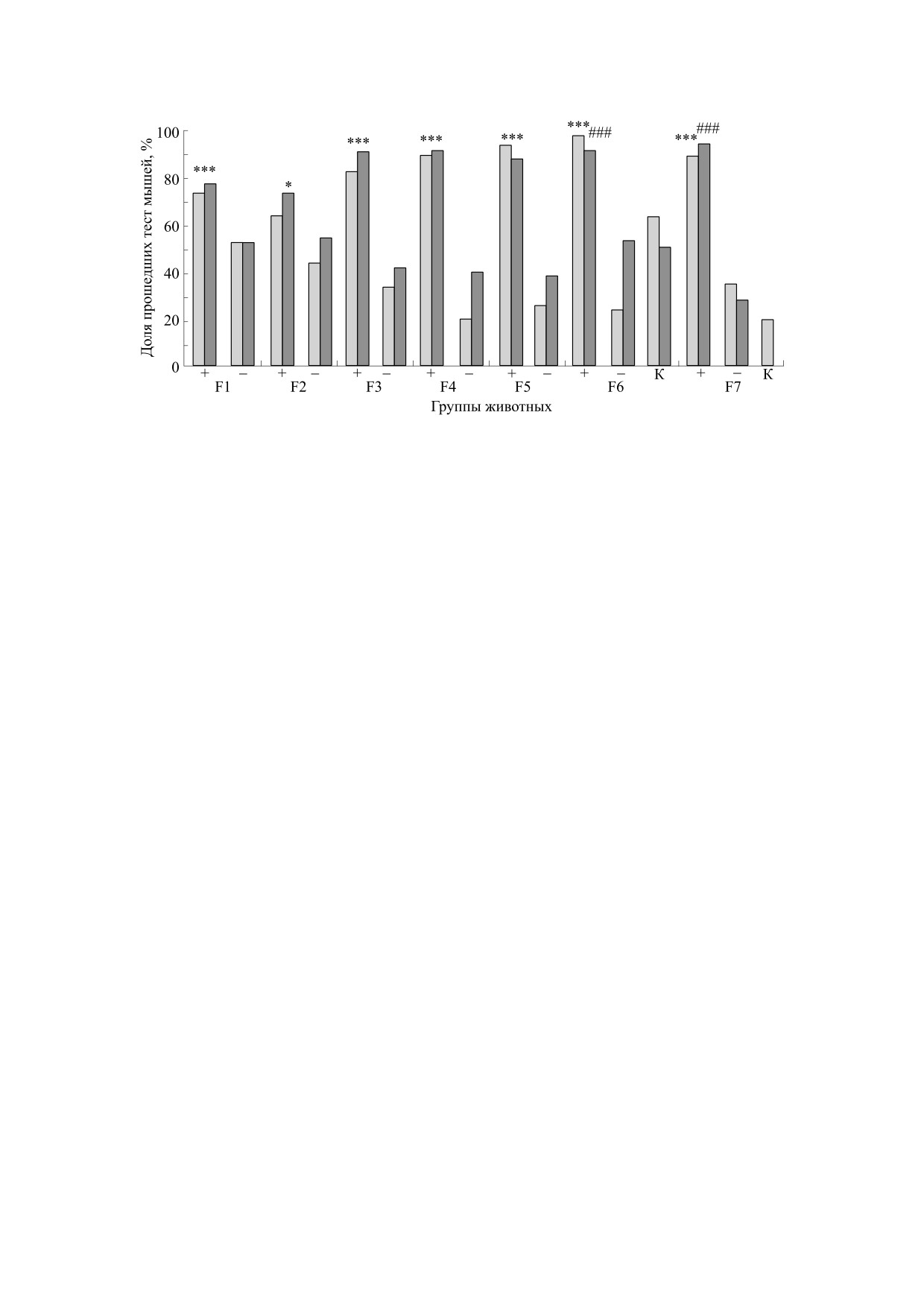
Рис. 5. Доли мышей (ось ординат, %) поколений селекции F1-F7 («+» и «-»), и мышей контрольной популяции («К»),
успешно решивших тест ПВУ (стадии 3 и 4 теста, светло-серые и темно-серые столбцы соответственно, когда лаз закрыт
пробкой); *** - достоверное отличие от доли линии «минус» при р < 0.001, ### - достоверное отличие от группы «К», р <
0.001 (φ-тест Фишера для оценки достоверности различий альтернативных долей).
время среди контрольных мышей КоЭКС (тести-
ОБСУЖДЕНИЕ
рованных в те же сроки, что и мыши F3), «не ма-
нипулировавших» пробкой животных было 8 из
Генетические основы решения животными те-
стов на элементарную рассудочную деятельность
34. Различия в успешности решения «трудных»
(на примере теста на экстраполяцию) анализиро-
этапов теста ПВУ, выраженные достаточно четко,
вались Л.В. Крушинским и его коллегами ранее
начиная с F3, могут служить свидетельством эф-
[14, 29]. Это были сравнительные исследования
фективности отбора по этим контрастным при-
выполнения теста на экстраполяцию серебристо-
знакам. Следует напомнить, что обе сублинии
черными лисицами разных генотипов (на базе
происходили из линии ЭКС, в 20 поколениях ко-
ИЦиГ в Новосибирске), а также попытка селек-
торой проводили отбор на успешное решение ко-
ции крыс на способность к экстраполяции на ос-
гнитивного теста на экстраполяцию направления
нове популяции гибридов диких крыс-пасюков и
движения [27]. Сложности экспериментов такого
крыс лабораторной линии Крушинского-Мо-
рода известны [28, 29].
лодкиной. Селекция крыс была прекращена на
уровне F4, поскольку практически все животные
Сравнение времени решения теста при первом
этого поколения обнаружили столь высокий уро-
и втором предъявлениях теста с лазом, замаски-
вень тревожности в обстановке эксперимента,
рованном пробкой позволило увидеть дополни-
что определить их способность к решению теста
тельно различия между линиями «плюс» и «ми-
на экстраполяцию не представлялось возмож-
нус». Доли мышей, у которых латентный период
ным. Ввиду этого селекция мышей на способ-
решения второго предъявления пробки был коро-
ность к решению теста на экстраполяцию (т.е. со-
че, чем при первом ее предъявлении, в линии
здание линии ЭКС), начатая значительно позд-
«плюс» были больше, чем в линии «минус»
нее, проводилась с отбором производителей не
(рис. 6). Это можно интерпретировать как более
только по высоким показателям решения теста на
четкое проявление краткосрочной рабочей памя-
экстраполяцию, но и по проявлению тревожно-
ти у мышей линии «плюс», что находится в соот-
сти в обстановке этого эксперимента. Иными
ветствии с их более успешным решением теста
словами, в селекцию не брали животных, кото-
ПВУ на стадии предъявления лаза, закрытого
рые медленно и очень осторожно пили молоко
«пробкой».
через центральное отверстие (откуда начиналось
движение поилки с молоком, которую следовало
Тест на внимание с движущимися предметами. В
отыскать, перемещаясь в направлении ее исчез-
решении этого теста (впервые использованного в
новения). Для формирования производителей
данной работе) также выявились межлинейные
для получения следующего поколения мышей
различия, хотя следует отметить, что они были
линии ЭКС не брали также тех животных, кото-
более четкими в шестом поколении селекции (см.
рые обнаруживали признаки испуга при начале
рис. 7).
движения поилки, когда животное отскакивало
БИОФИЗИКА том 68
№ 5
2023
СЛОЖНАЯ СИСТЕМА МОЗГА МЫШИ
1053
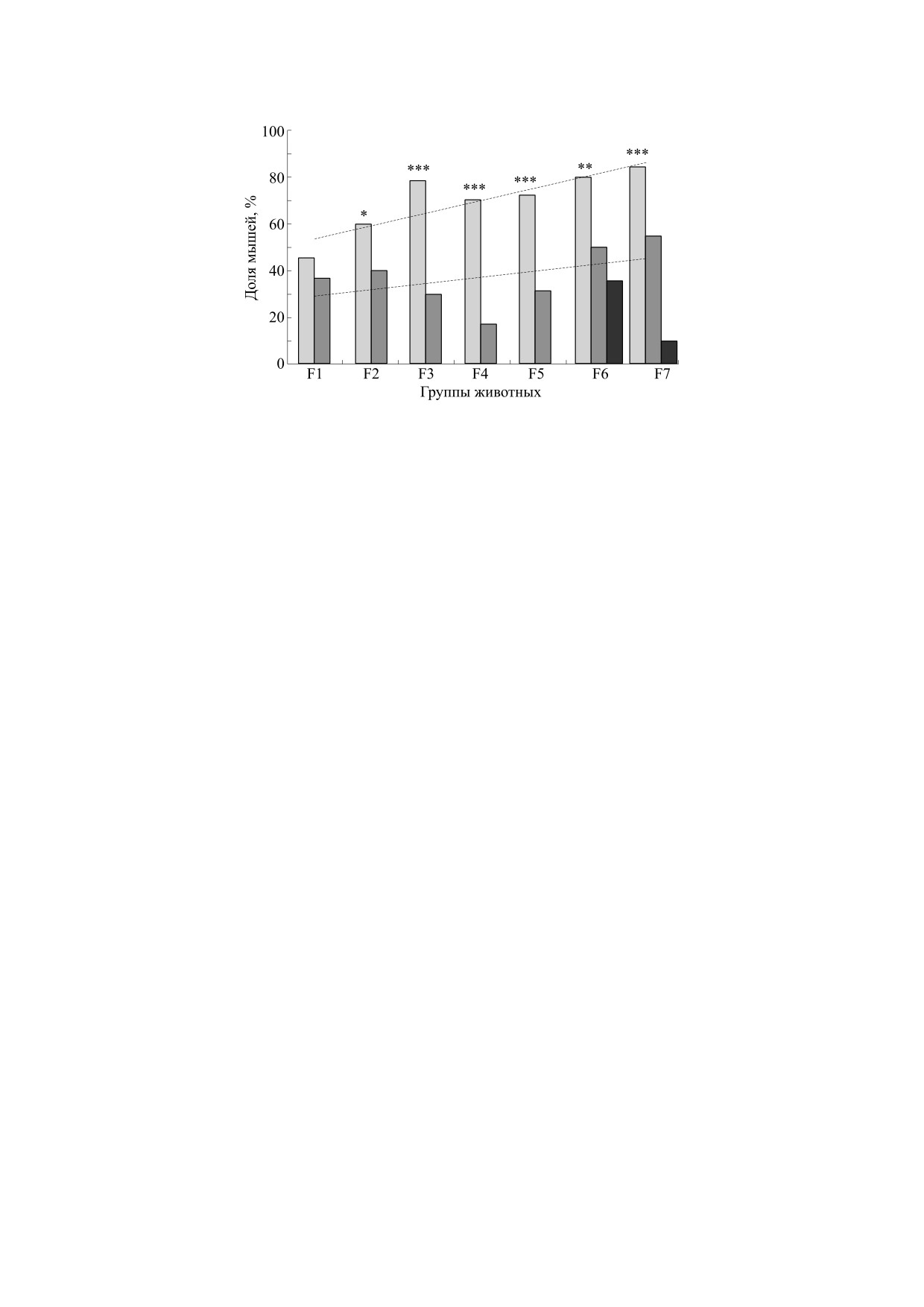
Рис. 6. Доли мышей ряда поколений селекции и контрольных мышей КоЭКС групп, у которых латентный период
решения теста на поиск входа в укрытие при втором предъявлении лаза, закрытого пробкой, стал короче, чем при первом
предъявлении этой задачи. Светло-серые столбики - мыши линии «плюс», темно-серые столбики - мыши линии
«минус», черные столбики - мыши контрольной неселектированной популяции КоЭКС; *, **, *** - достоверное
отличие от показателей групп линии «минус» и контрольной популяции при р < 0.05, 0.01 и 0.001 (φ-тест Фишера для
оценки достоверности различий альтернативных долей).
от центрального отверстия, не успев проследить
«плюс» и «минус» обладали в определенной сте-
начало движения поилки [26].
пени «пониманием» принципа неисчезаемости,
что и проявилось в том, что подавляющее боль-
У мышей первых поколений селекции линии
шинство мышей, даже не решивших стадии теста
ЭКС было отмечено некоторое превышение доли
с пробками, пытались проникнуть в лаз, толкая и
правильных решений теста на экстраполяцию
приподнимая пробку. Можно сделать заключе-
над случайным 50%-м уровнем [26], однако в по-
следующих поколениях (начиная с F9-F10) такое
ние, что селекция на успешное и не успешное ре-
превышение стало обнаруживаться нерегулярно.
шение теста ПВУ в его варианте с пробкой вы-
Можно полагать, что селекция на способность к
явила различия не только в решении теста, но и в
экстраполяции не была успешной (и у крыс, и у
способности животного «добиться» конкретного
мышей) из-за сложности этого признака [25, 26].
решения задачи. Это обстоятельство заставляет
Мы полагаем также, что отбор на этот когнитив-
предположить, что различия между новыми ли-
ный признак мог сопровождаться усилением
ниями затронули проявление так называемых
функции глутаматергической системы в ЦНС,
«исполнительных функций» (executive function)
поскольку роль этой системы в обеспечении пла-
[20, 25, 35].
стических изменений поведения была показана в
большом числе исследований [30-32]. Однако
Успешность выполнения теста, т.е. реализа-
при отборе на высокие показатели решения дан-
ция «исполнительных функций» у мышей разно-
ного теста изменения в глутаматергической си-
го генотипа была показана в одной из первых ра-
стеме могли проявиться не только в структурах
бот с использованием данного теста [21]. При вы-
переднего мозга, важных для проявления когни-
полнении теста ПВУ при открытом лазе, а также
тивных способностей, но и в других структурах.
при лазе, замаскированном стружкой, различия в
Существуют свидетельства участия в реализации
реакцией страха и тревоги глутаматных рецепто-
проявлении исполнительных функций обнару-
ров нейронов дорсальной части центрального се-
живались не в успешности решения этих этапов,
рого вещества (см., например, работы [33, 34]).
а, видимо, только в быстроте реализации реше-
ния. Мыши «плюс»-групп переходили в темноту с
Логическая структура теста и на экстраполя-
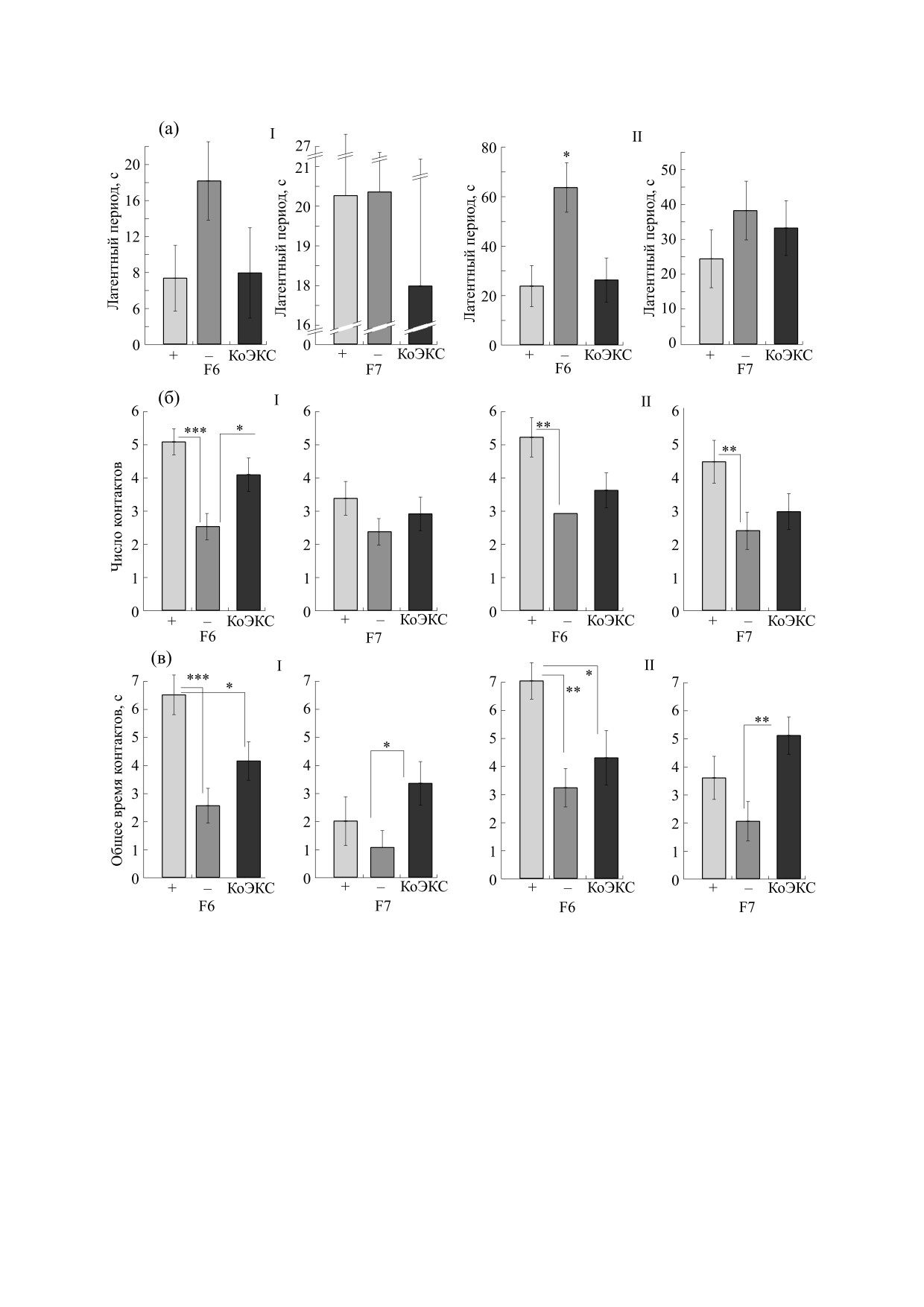
меньшим латентным периодом (рис. 4). С целью
цию, и на ПВУ предполагает, что для успешного
предварительной оценки возможных межлиней-
решения животное должно оперировать законо-
ных различий в экспрессии именно «исполни-
мерностью «неисчезаемости», т.е. понимать, что
тельных функций» мышам двух поколений селек-
исчезнувший (из поля зрения) объект (поилка с
молоком или замаскированный лаз) продолжают
ции (F6 и F7) был предъявлен тест на внимание,
существовать, и их можно искать. Данные насто-
т.е. на прослеживание движения предмета (не ис-
ящей работы показывают, что мыши линий
чезавшим из поля зрения).
БИОФИЗИКА том 68
№ 5
2023
1054
ПОЛЕТАЕВА и др.
Рис. 7. Реакции мышей-самцов (средние значения ± ошибка среднего) групп F6 и F7 на движущиеся в поле зрения
новые предметы (тест на внимание): (а) - латентный период первого подхода к предмету, (б) - число контактов с
предметом за 40 с движения предмета по кругу, (в) - общее время контактов с предметом. I - Предмет, предъявленный
первым, II - предмет, предъявленный последним; светло серые столбики - линия «плюс», темно серые столбики -
линия «минус», черные столбики - контрольная популяция КоЭКС. *, **, *** - достоверные различия при р < 0.05, 0.01
и 0.001 соответственно (однофакторный анализ ANOVA, фактор «генотип», post hoc LSD-тест по Фишеру).
ЗАКЛЮЧЕНИЕ
следования нет. В настоящее время основное
внимание нейрогенетиков, занимает анализ мо-
Следует отметить, что в литературе по анализу
дуляции экспрессии большого числа генов у гене-
когнитивного поведения лабораторных грызунов
(т.е. их способности к решению элементарных
тически измененных животных, и подобный спи-
логических задач) аналогов представленного ис-
сок работ насчитывает многие десятки исследо-
БИОФИЗИКА том 68
№ 5
2023
СЛОЖНАЯ СИСТЕМА МОЗГА МЫШИ
1055
ваний. Особенно много получено данных,
6. A. Fernández-Blanco and M. Dierssen, Int. J. Mol.
принципиально важных для исследования генеза
Sci., 21, 9039 (2020). DOI: 10.3390/ijms21239039
заболеваний ЦНС человека (например, работы
7. M. Mameli and P. Bateson, Phil. Trans. R. Soc. B, 366,
[35-37] и др.). Но селекционные эксперименты, в
436 (2011). DOI: 10.1098/rstb.2010.0174
которых проводился бы отбор по экспрессии ко-
8. A. Manning, In: Psychobiology: Biological basis of be-
гнитивных признаков, практически не проводят-
havior, Ed. by J. McGough (Acad Press, N-Y, London,
ся, хотя экспериментальный подход с использо-
1971), pp. 1-52.
ванием «батареи тестов» [20, 38-40] используется
9. W. C. Rothenbuhler, Am. Zool., 4, 111 (1964)
уже достаточно давно. В целом эта область нейро-
10. И. И. Полетаева, Природа, № 8, 150 (1999).
биологии, несмотря на большой прогресс в выяв-
11. R. W. Williams, R. C. Strom, and D. Goldowitz, J.
лении важных для когнитивного поведения
Neurosci., 18 (1), 138 (1998).
структур мозга, многих сигнальных путей и групп
12. P. Bateson and M. Mameli, Dev. Psychobiol., 49 (8),
специфических нейронов, остается пока недоста-
818 (2007). DOI: 10.1002/dev.20277
точно исследованной. Оценивая различия в ре-
13. B. Anderson, Am. J. Ment. Retard., 99 (1), 50 (1994).
зультатах анализа поведения лабораторных гры-
14. Л. В. Крушинский, Элементарная рассудочная дея-
зунов, точнее, его аспектов, сходных с темой на-
тельность (Изд-во МГУ, М., 1976).
стоящей работы, следует также помнить о
15. I. I. Poletaeva and Z. A. Zorina, Russ. J. Cogn. Sci., 1
большом числе часто неконтролируемых пере-
(3), 31 (2014).
менных, которые могут влиять даже на «знак» по-
16. A. R. Jensen, Ann. N. Y. Acad. Sci., 702 (1), 103
лучаемых межлинейных или межгрупповых раз-
(1993). DOI: 10.1111/j.1749-6632.1993.tb17244.x
личий (важность соблюдения единой последова-
17. B. Anderson, In Novartis Found Symp., discussion 90-5,
тельности тестов, учитывать влияния признаков
233, 79 (2000).
конкретного экспериментатора, например, в ра-
боте [28], и др.).
18. L. D. Matzel and S. Kolata, Neurosci. Biobehav. Rev.,
34
(1),
23
(2010).
DOI:
10.1016/j.neubior-
Таким образом, в работе описаны достоверные
ev.2009.07.002
различия в решении элементарной логической
19. S. Kolata, K. Light, H. C. Grossman, et al., Learn.
задачи, т.е. различия в экспрессии «исполнитель-
Mem., 14 (1), 22 (2007). DOI: 10.1101/lm.408507
ных функций» у мышей, которые прошли семь
20. M. J. Galsworthy, J. L. Paya-Cano, L. Liu, et al., Be-
поколений селекции на «успешность» и «не-
hav. Genet., 35 (5), 675 (2005). DOI: 10.1007/s10519-
успешность» решения теста на «неисчезаемость».
005-3423-9
21. N. M. Ben Abdallah, J. Fuss, M. Trusel, et al., Exp.
ФИНАНСИРОВАНИЕ РАБОТЫ
Neurol. 227 (1), 42 (2011). DOI: 10.1016/j.expneu-
rol.2010.09.008
Работа выполнена в рамках Государственной
22. S. Kolata, K. Light, and L. D. Matzel, Intelligence, 36,
программы № ААА-А16-116021660055-1.
619 (2008), DOI: 10.1016/j.intell.2007.12.001
23. S. R. O. Nilsson, J. Alsiöa,. E. M. Somervillec, and
P. G. Clifton, Neurosci. Biobehav. Rev., 56, 1 (2015).
КОНФЛИКТ ИНТЕРЕСОВ
DOI: 10.1016/j.neubiorev.2015.06.015
Авторы заявляют об отсутствии конфликта
24. B. Kolisnyk, M. A. Al-Onaizi,. P. H. F. Hirata, et al., J.
интересов.
Neurosci., 33 (37), 14908 (2013).
25. О. В. Перепелкина и И. И. Полетаева, Докл. РАН,
499, 322 (2021).
СПИСОК ЛИТЕРАТУРЫ
26. О. В. Перепелкина, Н. В. Маркина, В. А. Голибро-
1. S. M. Tomchik and R. L. Davis, Nature, 453 (7199),
до и др., Журн. высш. нерв деятельности им. И.П.
1192 (2008).
Павлова, 61 (6),742 (2011).
27. K. S. Van Driel and J. C. Talling, Behav. Brain Res.,
2.
2Z. Reznikova, Anim. Cogn., 23 (6), 1143- (2020).
159 (2), 243 (2005). DOI: 10.1016/j.bbr.2004.11.005
DOI: 10.1007/s10071-020-01423-x
28. P. Georgiou, T. M. Zanos, X. Mou, et al., Nat. Neuro-
3. Behavioral Genetics of the Mouse, Ed. by W. E. Crusio,
sci., 25 (9), 1191 (2022). DOI: 10.1038/s41593-022-
F. Sluyter, R. T. Gerlai, and S. Pietropaolo, Genetics of
01146-x
Behavioral Phenotypes (Cambridge University Press,
29. И. И. Полетаева и З. А. Зорина, в сб. Формирование
2013), v. 1, pp. 357.
поведения животных в норме и патологии: к 100-ле-
4. А. Зорина и А. А. Смирнова, О чем рассказали «го-
тию со дня рождения Л. В. Крушинского (1911-
ворящие обезьяны. Способны ли высшие животные
1984), Сост. И. И. Полетаева и З. А. Зорина (ЯСК,
оперировать символами? (Изд-во «Языки славян-
М., 2013), сс. 2-8.
ской культуры», 2008).
30. R. M. O'Connor, B. C. Finger, P. J. Flor, and J. F. Cry-
5. E. I. Rogaev, Front. Genet., 3 (3), ID 00045 (2012).
an, Eur. J. Pharmacol., 639 (1-3), 123 (2010). DOI:
DOI: 10.3389/fgene.2012.00045
10.1016/j.ejphar.2010.02.059
БИОФИЗИКА том 68
№ 5
2023
1056
ПОЛЕТАЕВА и др.
31. J. L. Brigman, E. M. Powell, G. Mittleman, and
36. L. Giménez-Llort, S. N. Schiffmann, T. Shmidt, et al.,
J. W. Young, Physiol. Behav., 107 (5), 666 (2012). DOI:
Neurobiol. Learn. Mem., 87 (1), 42 (2007). DOI:
10.1016/j.physbeh.2011.12.024
10.1016/j.nlm.2006.05.004
37. Q. Li, S. Y. Chan, K. K. Wong, et al., Behav. Genet., 46
32. D. A. Hamilton and J. L. Brigman, Genes, Brain and
(4), 529-537 (2016).
Behavior, 14, 4 (2015).
38. J. R. Homberg, E. J. Kyzar, M. Nguyen, et al., Neuro-
33. A. E. Reimer, A. R. De Oliveira, and M. L. Brandao,
sci. Biobehav. Rev.,
65,
292
(2016). DOI:
Neuroscience, 219, 72 (2012).
10.1016/j.neubiorev.2016.03.013
39. C. Locurto, A. Benoit, C. Crowley, and A. Miele, J.
34. R. R. Rozeske, D. Jercog, N. Karalis, et al., Neuron, 97
Comp. Psychol., 120 (4), 378 (2006).
(4), 898 (2018). DOI: 10.1016/j.neuron.2017.12.044
40. H. M. Ene, N. Z. Kara., N. Barak, et al., Acta Neuro-
35. J. Talpos and M. Shoaib, Handbook Exp. Pharmacol.,
psychiatr.,
28
(2),
85
(2016). DOI:
10.1017/
228, 191 (2015). DOI: 10.1007/978-3-319-16522-6_6
neu.2015.53
A Complex System of the Mouse Brain - Selection for Successful Solution
of a Cognitive Test
I.I. Poletaeva*, O.V. Perepelkina*, and Z.A. Zorina*
*Faculty of Biology, Lomonosov Moscow State University, Leninskie Gory 1/12, Moscow, 119234 Russia
The study provides a review of experimental research on selection of mice that yields positive results. The se-
lection of mice was high (the task required mice to understand that an object which has become invisible, still
exists and could be found) and poor performance in a cognitive test. The results obtained revealed better
short term memory in mice, selected for high performance scores, and that these mice also showed higher
performance in tests for novelty and attention. This suggests that differences between mice used in the exper-
iment are associated not only with cognitive abilities in tests, but also with more pronounced “executive
functions”.
Keywords: animal cognitive abilities, elementary logic task, behavior genetics, selection, laboratory mice
БИОФИЗИКА том 68
№ 5
2023