БИОФИЗИКА, 2023, том 68, № 5, с. 1031-1037
БИОФИЗИКА CЛОЖНЫX CИCТЕМ
УДК 577.353
ПРОТЕОСТАЗ БЕЛКА ТЕПЛОВОГО ШОКА HSP90 В СКЕЛЕТНЫХ
МЫШЦАХ ДЛИННОХВОСТОГО СУСЛИКА В ПЕРИОД ГИБЕРНАЦИИ
© 2023 г. Ю.В. Грицына*, С.С. Попова*, Г.З. Михайлова*, Л.Г. Бобылёва*, С.Н. Удальцов**,
О.С. Моренков***, Н.М. Захарова***, И.М. Вихлянцев*, ****, #
* Институт теоретической и экспериментальной биофизики РАН,
142290, Пущино Московской области, Институтская ул., 3
**Институт физико-химических и биологических проблем почвоведения - обособленное подразделение ФИЦ
«Пущинский научный центр биологических исследований Российской академии наук»,
Институтская ул., 2, Пущино Московской области, 142290, Россия
***Институт биофизики клетки Российской академии наук - обособленное подразделение ФИЦ «Пущинский
научный центр биологических исследований Российской академии наук»,
Институтская ул., 3, Пущино Московской области, 142290, Россия
****Институт фундаментальной медицины и биологии Казанского федерального университета,
ул. Карла Маркса, 76, Казань, 420012, Россия
#E-mail: ivanvikhlyantsev@gmail.com
Поступила в редакцию 24.07.2023 г.
После доработки 01.08.2023 г.
Принята к публикации 02.08.2023 г.
Исследованы изменения содержания белка теплового шока 90 (HSP90) в m. soleus (содержит
преимущественно волокна, экспрессирующие «медленную» изоформу I ТЦМ) и m. gastrocnemius
(содержит преимущественно волокна, экспрессирующие «быстрые» изоформы II ТЦМ) истинного
гибернанта длиннохвостого суслика (Urocitellus undulatus) в разные периоды годового цикла -
летней активности (сезонный контроль), гипотермии/спячки, зимней (межбаутной) активности.
Обнаружено, что несмотря на развитие атрофических изменений, более выраженных в «быстрой»
m. gastrocnemius, содержание HSP90 в обеих мышцах не изменялось на протяжении всего периода
гибернации. Обсуждается роль HSP90 в поддержании стабильности молекул гигантского
саркомерного белка титина в периоды входа животного в состояние гипотермии и выхода из этого
состояния, когда увеличивается активность кальпаиновых протеаз вследствие повышенного
содержания Са2+ в цитозоле мышечных клеток, а также в период гипотермии, когда активность
кальпаинов, по всей вероятности, не ингибируется полностью. В период зимней/межбаутной
активности, когда наблюдается повышенный оборот титина в поперечно-полосатых мышцах
суслика, константное содержание HSP90, по-видимому, необходимо для правильного фолдинга
заново синтезированных молекул титина и их встраивания в саркомеры, а также для удаления
неправильно свернутых и отслуживших молекул/фрагментов титина и других белков. Таким
образом, протеостаз HSP90 в скелетных мышцах длиннохвостого суслика может вносить вклад в
поддержание стабильного уровня титина и, возможно, других саркомерных белков в период
гибернации, что, в свою очередь, будет способствовать поддержанию высокоупорядоченной
саркомерной структуры и необходимого уровня сократительной активности мышц в разные фазы
цикла «спячка-бодрствование».
Ключевые слова: гибернация, длиннохвостый суслик Urocitellus undulatus, скелетные мышцы, белки
теплового шока, HSP90.
DOI: 10.31857/S0006302923050241, EDN: NASQXV
денческих, физиологических, клеточных и моле-
Гибернация/зимняя спячка млекопитающих -
это состояние гипотермии и сниженного метабо-
кулярных стратегий, позволяющих млекопитаю-
лизма, в которое животное входит вследствие
щим выживать в суровых условиях окружающей
эволюционно закрепленной совокупности пове-
среды [1]. Многочисленные исследования меха-
низмов адаптации зимоспящих животных к усло-
виям зимней спячки свидетельствуют о наличии у
Сокращения: ТЦМ - тяжелые цепи миозина, HSP90 -
белок теплового шока 90.
разных видов гибернантов общих молекулярно-
1031
1032
ГРИЦЫНА и др.
генетических и других стратегий выживания в
ров в поперечно-полосатых мышцах длиннохво-
условиях гибернации. Так, проведенные за по-
стого суслика в периоды зимней активности [12].
следние годы исследования выявили дифферен-
В данной работе в двух скелетных мышцах
циально экспрессирующиеся гены в поперечно-
длиннохвостого суслика исследованы сезонные
полосатых мышцах, а также других органах зимо-
изменения содержания белка теплового шока 90
спящих, что свидетельствует о молекулярно-ге-
(HSP90), который, являясь шапероном и важным
нетических механизмах, регулирующих сезонные
компонентом механизма внутриклеточного
изменения активности многих сигнальных путей
«контроля качества белка» (protein quality control,
[2-5]. В научной литературе появились новые
PQC), играет также важную роль в регуляции ста-
термины: «гибернационный фенотип», «несуще-
бильности молекул гигантского саркомерного
ственные гены и процессы» (nonessential genes and
белка титина [13]. В научной литературе также об-
processes), которые «выключаются», чтобы сохра-
суждается, что поддержание стабильного уровня
нить энергоресурсы, необходимые для выжива-
или увеличение содержания белков теплового
ния в условиях зимней спячки [6].
шока, в том числе и HSP90, в разных тканях и ор-
Однако важно отметить, что несмотря на на-
ганах, включая мышцы, как у зимоспящих мле-
личие у разных зимоспящих животных (летучих
копитающих [14-18], так и у других животных
мышей, сонь, сусликов, медведей и др.) общих
(черепахи [19], лягушки [20]), впадающих в состо-
особенностей их «гибернационного фенотипа»
яние оцепенения и гипотермии, может являться
(например, «fast-to-slow» трансформации миози-
общей молекулярной стратегией, защищающей
на и мышечных волокон [7,8]; переключения ме-
клетки и макромолекулы от повреждений в усло-
таболизма с углеводного на липидный [9]), в пе-
виях стресса во время гипотермии и при выходе
риод гибернации у этих животных наблюдаются
из этого состояния.
неодинаковые и часто противоположные измене-
Принимая во внимание процитированные вы-
ния на молекулярном уровне. Эти данные позво-
ше данные о белках теплового шока, данные о
ляют предположить, что у каждого вида зимоспя-
«fast-to-slow» изменениях в скелетных мышцах
щего животного существует свой уникальный
разных видов зимоспящих в период гибернации
подход к «погружению» в состояние гибернации,
[7, 8], а также данные о преобладающем синтезе
что, несомненно, является чрезвычайно интерес-
титина в мышцах длиннохвостого суслика в пери-
ным научным аспектом, хотя это значительно
од зимней активности [12], мы ожидали получить
усложняет поиск уникальных белковых комплек-
следующие результаты: (1) обнаружить констант-
сов с целью использования молекулярно-клеточ-
ный уровень HSP90 в «медленной» мышце m. so-
ных стратегий гибернантов для лечения/предот-
leus на фоне отсутствия или слабо выраженных
вращения негативных изменений у человека или
атрофических изменений в период гипотер-
у негибернирующих животных. В связи с этим ис-
мии/спячки; (2) выявить уменьшение содержа-
следования особенностей «гибернационного фе-
ния HSP90 в «быстрой» мышце m. gastrocnemius
нотипа» длиннохвостого суслика (Urocitellus un-
на фоне более выраженных атрофических изме-
dulatus), являющегося наименее изученным среди
нений в период гипотермии/спячки; (3) обнару-
других видов зимоспящих, имеют большую науч-
жить восстановление уровня HSP90 в m. gastroc-
ную значимость.
nemius и возможное увеличение содержания это-
Исследования, проведенные нами ранее, вы-
го белка в m. soleus длиннохвостого суслика в
явили уникальную молекулярную стратегию
период зимней (межбаутной) активности.
длиннохвостого суслика, заключающуюся в под-
держании стабильного уровня гигантских мио-
ОБЪЕКТЫ И МЕТОДЫ
фибриллярных белков толстых и тонких нитей
титина и небулина в поперечно-полосатых мыш-
Эксперименты проводили на истинном гибер-
цах в течение всего гибернационного сезона [10-
нанте длиннохвостом суслике Urocitellus undulatus
12]. Так, во время оцепенения/спячки в сердеч-
(прежнее название Spermophilus undulatus). Жи-
ной и ряде скелетных мышц сусликов наблюда-
вотные были отловлены в окрестностях города
лось незначительное уменьшение (в ~1,2 раза) со-
Якутска и доставлены в виварий ИБК РАН (Пу-
держания титина. Во время межбаутной/зимней
щино, Московская область), где они содержались
активности наблюдался преимущественный син-
в индивидуальных клетках при естественном
тез титина, что приводило к быстрому восстанов-
освещении, температуре воздуха 20-21°С и влаж-
лению нормального содержания этого белка в
ности 65-70%. Гнездовой материал и пища - ad li-
мышцах [12]. При этом не наблюдалось наруше-
bitum. В экспериментах использовали животных
ний высокоупорядоченной структуры саркоме-
обоих полов массой 550-750 г. Животные были
БИОФИЗИКА том 68
№ 5
2023
ПРОТЕОСТАЗ БЕЛКА ТЕПЛОВОГО ШОКА HSP90
1033
разделены на три группы: «Летняя активность»
PVDF-мембрану. Были использованы первичные
(нормотермия, 38°С, июнь-июль, n = 6); «Гипо-
антитела к HSP90α/ß
(1:2000, номер клона
термия» (глубокое оцепенение/спячка, декабрь-
6H1/F8, получены в лаборатории проблем кле-
март, температура сердечной мышцы 1.5-2.0°C,
точного стресса (ИБК РАН, рук. лаб. О.С. Мо-
продолжительность периода гипотермии 5-11 су-
ренков), более подробная информация об этих
ток, n = 6); «Зимняя активность» (нормотер-
антителах представлена в работе [12]) и GAPDH
мия/межбаутная активность, 36-37°С, периоди-
(1:2000, ab37168, Abcam, Великобритания). В ка-
чески повторяющиеся временные промежутки
честве вторичных антител использовали антите-
(не более суток) между периодами гипотермии,
ла, конъюгированные с щелочной фосфатазой
взятие материала через 5-14 ч эутермного состо-
(1:3000, ab6722 и ab6790, Abcam, Великобрита-
янии после пробуждения, n = 6). Перед сезоном
ния). Для визуализации белковых полос на мем-
спячки сусликов переносили в темное помеще-
бранах использовали раствор NBT/BCIP (Roche,
ние с температурой 1-3°С и перед эксперимен-
Германия). Отмытые гели и блот-мембраны
том размещали в деревянных боксах размером
оцифровывали, а затем проводили денситомет-
20×20×25 см с гнездовым материалом. Для реги-
рический анализ с помощью компьютерной про-
страции состояния животных в дно деревянного
граммы Total Lab v1.11 (Newcastle Upon Tyne, Ве-
бокса были вмонтированы термодатчики. Темпе-
ликобритания).
ратура «подстилки» у животного, пребывающего
в состоянии гипотермии (спячки) была порядка
Для определения атрофических изменений
2-4°С, а при выходе животного из спячки
мышечных волокон методом 3D-реконструкции
температура дна ящика поднималась до 12-16°С.
были взяты фрагменты вышеуказанных двух ске-
летных мышц сусликов трех экспериментальных
Процедура мониторинга и отбора животных
групп - «Летняя активность» (n = 5) (сезонный
проводилась сотрудниками лаборатории меха-
контроль), «Гипотермия» (n = 5), «Зимняя актив-
низмов природного и искусственного гипобиоза
ность» (n = 4). Фрагменты мышц фиксировали в
(ИБК РАН, рук. лаб. Н.М. Захарова) и подробно
течение 16 ч при комнатной температуре, погру-
описана в работе [21]. Животных декапитировали
жая в раствор, содержащий 4% параформальдеги-
с помощью гильотины, немедленно вскрывали
да, 2.5% глутарового альдегида и 50 мМ сахарозы
грудную полость и измеряли температуру в обла-
сти сердца температурным датчиком RET2
в 0.1 М Na-какодилатном буфере (pH 7.4). Для
(Physitemp, США) с точностью до 0.1°С. Мышцы
постфиксации использовали 2%-й раствор четы-
m. soleus («медленная») и m. gastrocnemius media-
рехокиси осмия. После сушки в этаноле и ацето-
lis («быстрая») вырезали, замораживали в жидком
не ткани заливали эпоксидной смолой EMBed и
азоте и хранили при -75°С. Активные суслики
затем выдерживали в течение 48 ч при 60°C. Се-
подвергались анестезии Золетилом (Virbac Sante
рийные срезы толщиной 9 мкм получали из бло-
Animale, Carros, Франция) (4 мг/кг, внутримы-
ков, залитых в эпоксидную смолу на пирамитоме
шечно).
(LKB 11800, Швеция). Визуализацию проводили
на микроскопе NU-2E (Carl Zeiss, Германия, объ-
Для определения методом Вестерн-блоттинга
ектив E25× Planachromat). Панорамные изобра-
сезонных изменений содержания белка HSP90
жения серийных срезов были выровнены относи-
экстракцию белков из скелетных мышц сусликов
тельно друг друга с помощью компьютерной про-
проводили в лизирующем буфере, содержащем
граммы IGL Align sEM Align. Ориентирами
12 мM трис-HCl, 1.2% додецилсульфата натрия,
служили контуры внешнего перимизия. Контуры
5 мM ЭГTA, 10% глицерина, 2% β-меркаптоэта-
нола, 5 мкг/мл леупептина и ингибитора E64,
каждого мышечного волокна были построены пу-
pH 7.0. Экстракцию GAPDH (референсного бел-
тем ручной сегментации в программе IGL Trace
ка) проводили в этом же лизирующем буфере.
(версия 1.20b). Для калибровки был принят во
Белковые образцы нагревали при 95°С в течение
внимание коэффициент усадки мышечной ткани
трех-четырех минут. На дорожки в геле наносили
(10%), вызванный процедурами постфиксации.
одинаковые объемы белковых образцов, вырав-
Трехмерные изображения мышечных волокон
ненных по концентрации общего белка, которую
были сформированы с использованием про-
определяли методом Бредфорда по протоколу из-
граммного обеспечения IGL Trace. Количествен-
готовителя (ЗАО «Силекс», Россия), используя
ные параметры рассчитывали с помощью ком-
бычий сывороточный альбумин как стандарт.
мерческой программы Actify’s 3D View. Объемы
ДСН-электрофорез проводили в 9.5-10% полиа-
120 и 150 мышечных волокон (из расчета 30 воло-
криламидном геле по методу, описанному в рабо-
кон для каждого мышечного образца) были рас-
те [22]. Перенос белков с геля осуществляли на
считаны для группы «Зимняя активность» и для
БИОФИЗИКА том 68
№ 5
2023
1034
ГРИЦЫНА и др.
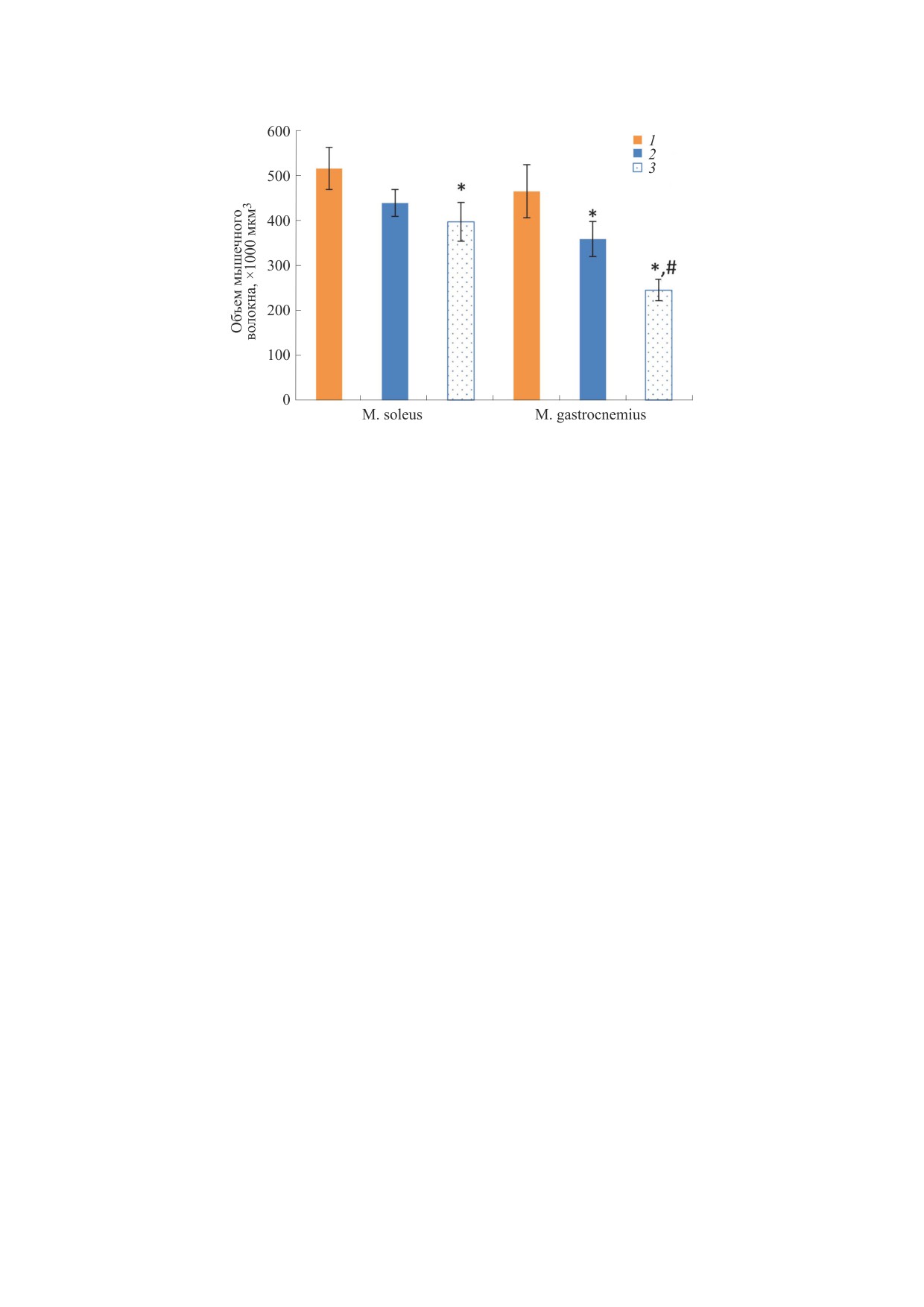
Рис. 1. Сезонные изменения объема мышечных волокон m. soleus и m. gastrocnemius длиннохвостого суслика: 1 -
«Летняя активность» (n = 5), 2 - «Гипотермия» (n = 5), 3 - «Зимняя активность» (n = 4). Значения представлены как M
± S D , г д е M - среднее значение, SD - стандартное отклонение; * - в сравнении с группой «Летняя активность», р <
0.05; # - в сравнении с группой «Гипотермия», р < 0.05.
групп «Летняя активность» и «Гипотермия» соот-
однако статистически значимых различий этого
ветственно.
параметра в сравнении с группой «Летняя актив-
ность» не было. Статистически значимое умень-
Статистическую обработку результатов иссле-
шение на 23% (p < 0.05) объема мышечных воло-
дования проводили при помощи пакета SigmaPlot
11.0 (Systat Software, США). В связи с тем, что рас-
кон m. soleus наблюдалось в группе «Зимняя ак-
пределение некоторых выборок данных не явля-
тивность». При этом статистически значимых
лось нормальным (согласно критерию Шапиро-
различий в атрофии мышечных волокон m. soleus
Уилка), для анализа достоверности наблюдаемых
между группами «Гипотермия» и «Зимняя актив-
различий использовали непараметрический од-
ность» не выявлено. Полученные результаты сви-
нофакторный дисперсионный анализ для по-
детельствуют о более выраженных атрофических
вторных измерений (односторонний дисперси-
изменениях в «быстрой» мышце m. gastrocnemius
онный анализ Краскела-Уоллиса) с последую-
и менее выраженной атрофии в «медленной»
щим парным сравнением по тесту Тьюки.
мышце m. soleus у длиннохвостого суслика в пе-
Различия считали статистически значимыми при
риод гибернации.
р < 0.05. Данные представлены как M ± SD, где
На рис. 2 представлены данные Вестерн-блот-
М - среднее значение, SD - стандартное откло-
анализа сезонных изменений содержания HSP90
нение.
в m. gastrocnemius и m. soleus длиннохвостого сус-
лика. Несмотря на развитие атрофических изме-
РЕЗУЛЬТАТЫ
нений (рис. 1), статистически значимых разли-
чий в содержании HSP90 в исследуемых скелет-
На рис. 1 представлены данные 3D-рекон-
ных мышцах сусликов трех экспериментальных
струкции объема мышечных волокон m. soleus и
групп не выявлено.
m. gastrocnemius сусликов трех эксперименталь-
ных групп. Результаты указывают на развитие
ОБСУЖДЕНИЕ РЕЗУЛЬТАТОВ
атрофических изменений в двух скелетных мыш-
цах длиннохвостого суслика в период гибернаци-
В целом наши предположения о поддержании
онного сезона. В частности, обнаружено умень-
константного уровня HSP90 в скелетных мышцах
шение на 23% (p < 0.05) объема мышечных воло-
длиннохвостого суслика в период гибернации
кон «быстрой» икроножной мышцы сусликов
подтвердились. В чем может заключаться физио-
группы «Гипотермия» и на 48% (p < 0.01) группы
логический смысл подобной молекулярной стра-
«Зимняя активность» в сравнении с группой
тегии? Белки теплового шока, в том числе и
«Летняя активность». Объем мышечных волокон
HSP90, являются важными компонентами меха-
«медленной» m. soleus сусликов группы «Гипо-
низма внутриклеточного «контроля качества бел-
термия» имел тенденцию к уменьшению на ~15%,
ка» [23]. Внутриклеточный «контроль качества
БИОФИЗИКА том 68
№ 5
2023
ПРОТЕОСТАЗ БЕЛКА ТЕПЛОВОГО ШОКА HSP90
1035
Рис. 2. Вестерн-блот анализ сезонных изменений содержания HSP90 в «быстрой» m. gastrocnemius (n = 6) и «медленной»
m. soleus (n = 6) мышцах длиннохвостого суслика: 1 - «Летняя активность»; 2 - «Гипотермия»; 3 - «Зимняя активность»;
GAPDH - референсный белок.
белка» необходим для поддержания баланса меж-
протеолитическим ферментом титина. Эти дан-
ду деградацией белка и его синтезом. Поддержа-
ные свидетельствуют о положительной роли
ние такого баланса особенно важно в «долгожи-
HSP90 в регуляции стабильности молекулы тити-
вущих», интенсивно работающих и испытываю-
на [13].
щих значительные механические нагрузки
Результаты, полученные в нашем исследова-
клетках поперечно-полосатых мышц. Большое
нии, свидетельствуют о константном содержании
количество белковых компонентов должно быть
HSP90 в течение всего гибернационного периода
синтезировано, подвергнуто правильному фол-
как в «быстрой», так и в «медленной» скелетных
дингу и встроено в саркомер, тогда как пропорци-
мышцах длиннохвостого суслика, несмотря на
ональная часть соответствующих белков должна
развитие в них атрофических изменений. Анало-
быть подвергнута деградации [23, 24]. Дисбаланс
гичные результаты, свидетельствующие о поддер-
белкового гомеостаза приводит к накоплению не-
жании константного уровня HSP90 в течение все-
правильно свернутых белков и их агрегатов в ци-
го сезона спячки в двух других поперечно-поло-
сатых мышцах (миокарде и m. longissimus dorsi)
тозоле, которые являются токсичными для клет-
длиннохвостого суслика, получены нами ранее
ки, что, в конечном итоге, ведет к развитию мио-
[12]. Подобный протеостаз HSP90 несомненно
патии [24, 25]. Одним из «сложных» объектов для
будет играть положительную роль в поддержании
механизма внутриклеточного «контроля качества
стабильности молекул титина как в периоды вхо-
белка» является гигантский мышечный белок ти-
да животного в состояние гипотермии и выхода
тин. На сегодняшний день точные механизмы
из этого состояния, когда увеличивается актив-
«контроля качества» титина ни в норме, ни при
ность кальпаиновых протеаз вследствие повы-
патологии, ни при гибернации до конца неясны.
шенного содержания Са2+ в цитозоле мышечных
Шаперон HSP90 широко распространен во
клеток (см. для ссылок работу [12]), так и в период
всех тканях живых рганизмов и принимает уча-
гипотермии, когда активность кальпаинов, судя
стие в фолдинге, поддержании структуры белков,
по нашим данным, не ингибируется полностью
их деградации, в предотвращении белковой агре-
[12, 27]. В период зимней/межбаутной активно-
гации, а также в процессах внутриклеточной сиг-
сти, когда наблюдается повышенный оборот
нализации [26]. В саркомере HSP90 связывается с
(turnover) титина в поперечно-полосатых мыш-
миозином, однако в комплексе с белками SET-се-
цах длиннохвостого суслика [12], константное со-
мейства (содержат SET-домен, обладающий ме-
держание HSP90 будет необходимо для правиль-
тилтрансферазной активностью) HSP90 спосо-
ного фолдинга заново синтезированных молекул
бен связываться с N2A-доменом титина, защи-
титина и их встраивания в саркомеры, а также для
щая тем самым от протеолиза эту часть молекулы
удаления неправильно свернутых и «старых» мо-
титина в I-зоне саркомера. Для взаимодействия с
лекул/фрагментов титина и других белков. Таким
титином HSP90 должен быть метилирован с по-
образом, протеостаз HSP90 в скелетных мышцах
мощью SMYD2 [13]. После глутатионилирования
длиннохвостого суслика может вносить вклад в
или окисления SMYD2 этот комплекс диссоции-
поддержание стабильного уровня титина и других
рует и N2A-область титина оказывается доступ-
саркомерных белков в период гибернации, что, в
ной для расщепления кальпаином-1 - основным
свою очередь, будет способствовать поддержа-
БИОФИЗИКА том 68
№ 5
2023
1036
ГРИЦЫНА и др.
нию высокоупорядоченной саркомерной струк-
СОБЛЮДЕНИЕ ЭТИЧЕСКИХ СТАНДАРТОВ
туры и необходимого уровня сократительной ак-
Все процедуры с животными проводили в со-
тивности мышц в разные фазы цикла «спячка-
ответствии с требованиями Комиссии по биоэти-
бодрствование»: вход в спячку, оцепенение, вы-
ке ИБК РАН и Европейской конвенции по защи-
ход из спячки, межбаутная активность.
те позвоночных животных, используемых для
экспериментальных и других научных целей (Eu-
Наши данные согласуются с результатами дру-
ropean
Communities
Council
Directive
гих исследований, тоже свидетельствующими о
(86/609/EEC)).
константом содержании разных белков теплового
шока в миокарде тринадцатиполосного суслика
(Ictidomys tridecemlineatus) в период гибернации
СПИСОК ЛИТЕРАТУРЫ
[18]. Авторами сделан вывод о цитопротекторной
1.
S. M. Mohr, S. N. Bagriantsev, and E. O. Gracheva,
роли белков теплового шока, способствующих
Annu. Rev. Cell Dev. Biol., 36, 315 (2020).
повышению устойчивости миокарда к стрессо-
2.
K. L. Vermillion, K. J. Anderson, M. Hampton, and
вым воздействиям в это время. Другие исследова-
M. T. Andrews, Physiol. Genomics, 47 (3), 58 (2015).
тели показали увеличение содержания HSP70 (в
3.
D. A. Mugahid, T. G. Sengul, X. You, et al., Sci. Rep.,
1.7 раза) в грудной мышце и двуглавой мышце
9 (1), 19976 (2019).
плеча у летучей мыши Murina leucogaster во время
4.
A. V. Goropashnaya, B. M. Barnes, and V. B. Fedorov,
спячки [15]. Был сделан вывод о важной роли по-
Sci. Rep., 10 (1):9010 (2020).
вышенного содержания HSP70 в предотвраще-
5.
E. Tseng, J. G. Underwood, B. D. Evans Hutzenbiler,
нии атрофии и поддержании нормальной сокра-
et al., Genes, Genomes, Genetics, 12 (3), jkab422
тительной способности скелетных мышц после
(2022).
6.
W. A. Ingelson-Filpula and K. B. Storey, Epigenomes,
их продолжительного (в течение трех месяцев)
2021 5(4):28 (2021).
неиспользования.
7.
C. J. Cotton, J. Exp. Biol., 219 (Pt 2), 226 (2016).
Мы полагем, что поддержание константного
8.
M. V. Lazareva, K. O. Trapeznikova, I. M. Vikhliant-
уровня HSP90 в скелетных мышцах длиннохво-
sev, et al., Biofizika, 57 (6), 982 (2012).
стого суслика вносит вклад в уменьшение протео-
9.
A. G. Hindle, A. Karimpour-Fard, L. E. Epperson,
et al., Am. J. Physiol. Regul. Integr. Comp. Physiol.,
лиза титина в период оцепенения, а также спо-
301 (5), 1440 (2011).
собствует быстрому обновлению и восстановле-
10.
I. M. Vikhlyantsev, E. V. Karaduleva, and Z. A. Pod-
нию нормального содержания титина в период
lubnaya, Biophysics, 53, 598 (2008).
межбаутных пробуждений. В совокупности полу-
11.
I. M. Vikhlyantsev and Z. A. Podlubnaya, Biochemis-
ченные нами ранее данные [10-12] и результаты
try (Moscow), 77 (13), 1515 (2012).
этой работы свидетельствуют о протеостазе тити-
12.
S. Popova, A. Ulanova, Y. Gritsyna, et al., Sci. Rep., 10
на и HSP90 в поперечно-полосатых мышцах
(1), 15185 (2020).
длиннохвостого суслика. Подобная молекуляр-
13.
L. T. Donlin, C. Andresen, S. Just, et al., Genes Dev.,
ная стратегия направлена на сохранение высоко-
26 (2), 114 (2012).
упорядоченной саркомерной структуры и под-
14.
S. F. Eddy, J. D. McNally, and K. B. Storey, Arch. Bio-
держание необходимого уровня сократительной
chem. Biophys., 435 (1), 103 (2005).
ативности мышц в разные фазы цикла «спячка-
15.
K. Lee, J. Y. Park, W. Yoo, et al., J. Cell Biochem., 104
бодрствование».
(2):642 (2008).
16.
C. W. Wu, K. K. Biggar, J. Zhang, et al., Genomics,
БЛАГОДАРНОСТИ
Proteomics & Bioinformatics, 13 (2), 119 (2015).
17.
B. E. Luu, S. Wijenayake, J. Zhang, et al., Comp. Bio-
Работа выполнена с использованием
chem. Physiol. B: Biochem. Mol Biol., 224, 26 (2018).
оборудования Сектора оптической микроскопии
18.
C. L. Childers, S. N. Tessier, and K. B. Storey, Peer J.,
и спектрофотометрии ЦКП ПНЦБИ РАН
7, e7587 (2019).
19.
A. Krivoruchko and K. B. Storey, J. Comp. Physiol. B,
180 (3), 403 (2010).
20.
H. M. Rabeae, S. S. Mahfouz, A. K. M. Abdel Latif,
ФИНАНСИРОВАНИЕ РАБОТЫ
et al., J. Therm. Biol. 114, 103490 (2023).
21.
Н.М. Захарова, Фундаментальные исследования,
Работа выполнена в рамках Государственного
6, 1401 (2014).
задания № 075-01025-23-01.
22.
U. K. Laemmli, Nature, 227 (5259), 680 (1970).
23.
S. Kötter and M. Krüger, Front. Physiol., 13, 914296
(2022).
КОНФЛИКТ ИНТЕРЕСОВ
24.
K. Hnia, T. Clausen, and C. Moog-Lutz, Trends Mol.
Med., 25 (9), 760 (2019).
Авторы заявляют об отсутствии конфликта
25.
Z. V. Wang and J. A. Hill, Cell Metab. 21(2), 215
интересов.
(2015).
БИОФИЗИКА том 68
№ 5
2023
ПРОТЕОСТАЗ БЕЛКА ТЕПЛОВОГО ШОКА HSP90
1037
26. P. Csermely, T. Schnaider, C. Soti, et al., Pharmacol.
27. S. S. Popova, I. M. Vikhlyantsev, N. M. Zakharova,
Ther., 79 (2), 129 (1998).
et al., Dokl. Biochem. Biophys. 472 (1), 56 (2017).
Proteostasis of Heat Shock Protein 90 in Skeletal Muscles of the Long-Tailed Ground
Squirrel during Hibernation
Yu.V. Gritsyna*, S.S. Popova*, G.Z. Mikhailova*, L.G. Bobyleva*, S.N. Udaltsov**,
O.S. Morenkov***, N.M. Zakharova***, and I.M. Vikhlyantsev*, ****
*Institute of Theoretical and Experimental Biophysics, Russian Academy of Sciences,
Institutskaya ul. 3, Pushchino, Moscow Region, 142290 Russia
**Institute of Physicochemical and Biological Problems of Soil Science, Russian Academy of Sciences,
Institutskaya ul. 2, Pushchino, Moscow Region, 142290 Russia
***Institute of Cell Biophysics, Russian Academy of Sciences, Institutskaya ul. 3, Pushchino, Moscow Region, 142290 Russia
****Institute of Fundamental Medicine and Biology, Kazan Federal University, ul. Karla Marksa 76, Kazan, 420012 Russia
We investigated changes in the content of heat shock protein 90 in m. soleus (comprised of mainly fibers ex-
pressing the MyHC slow isoform I) and m. gastrocnemius (composed of mainly fibers expressing the MyHC
fast isoforms II) of the long-tailed ground squirrel Urocitellus undulatus in different periods of the annual cy-
cle: summer activity (seasonal control), hypothermia/torpor, winter (interbout) activity. The content of the
protein in both muscles was found not to change throughout the entire hibernation period despite the devel-
opment of atrophic changes, more pronounced in fast m. gastrocnemius. The role of HSP90 in maintaining
the stability of giant sarcomeric titin protein molecules is discussed with reference to animal's entry into and
exit from hypothermia, when the activity of calpain proteases increases due to the increased content of Ca2+
in the cytosol of muscle cells; and with respect to the torpor, when the activity of calpains is, most likely, not
inhibited completely. During the interbout activity with an observed increased titin turnover in squirrel’s stri-
ated muscles, a constant content of HSP90 appears to be required for the correct folding of newly synthesized
titin molecules and their integration into sarcomeres, as well as for the removal of misfolded titin molecules
and other proteins. Thus, HSP90 proteostasis in skeletal muscles of the long-tailed ground squirrel can con-
tribute to maintaining a steady-state level of titin and, possibly, other sarcomeric proteins during hibernation,
which, in turn, will contribute to maintaining a highly ordered sarcomeric structure and the necessary level
of muscle contractile activity in different phases of the torpor-arousal cycle.
Keywords: hibernation, long-tailed ground squirrel Urocitellus undulatus, skeletal muscles, heat shock proteins,
HSP90
БИОФИЗИКА том 68
№ 5
2023