БИОФИЗИКА, 2023, том 68, № 5, с. 941-948
БИОФИЗИКА КЛЕТКИ
УДК 591.111.7; 576.524
ВЛИЯНИЕ ТАКСИФОЛИНА, КОНЪЮГАТА ТАКСИФОЛИНА
С ГЛИОКСАЛЕВОЙ КИСЛОТОЙ И НАРИНГЕНИНА
НА ФУНКЦИОНАЛЬНУЮ АКТИВНОСТЬ НЕЙТРОФИЛОВ
© 2023 г. В.С. Шубина*, #, М. И. Кобякова*, Ю.В. Шаталин*
*Институт теоретической и экспериментальной биофизики РАН,
Институтская ул., 3, Пущино Московской обл., 142290, Россия
#E-mail: shubinavictoria@yandex.ru
Поступила в редакцию 21.08.2023 г.
После доработки 28.08.2023 г.
Принята к публикации 06.09.2023 г.
Целью работы являлось изучение влияния таксифолина, конъюгата таксифолина с глиоксалевой
кислотой и нарингенина на фагоцитоз латексных частиц нейтрофилами и адгезивные свойства
данных клеток. Было показано, что таксифолин не влияет на фагоцитарную активность и
адгезивные свойства нейтрофилов. Конъюгат таксифолина с глиоксалевой кислотой значительно
ингибирует фагоцитарную активность нейтрофилов. В то же время, конъюгат значительно
усиливает адгезию данных клеток. Нарингенин снижает фагоцитарную активность нейтрофилов,
однако, в меньшей степени, чем конъюгат. Нарингенин также ингибирует адгезию нейтрофилов.
Полученные данные свидетельствуют о том, что полифенолы могут влиять на функциональную
активность нейтрофилов, что может играть важную роль в модуляции воспалительного процесса, в
частности когда пролонгированная активация нейтрофилов приводит к повреждению клеток и
тканей собственного организма.
Ключевые слова: фагоцитоз, адгезия, нейтрофилы, полифенолы.
DOI: 10.31857/S0006302923050149, EDN: PHHMYB
точная продукция АФК нейтрофилами может
Нейтрофилы являются наиболее многочис-
приводить к повреждению клеток и тканей соб-
ленной фракцией лейкоцитов, циркулирующих в
ственного организма, способствовать хрониза-
кровеносном русле. Данные клетки одними из
ции воспаления [2-4]. Избыточное образование
первых направляются к очагам инфекции и вос-
NET также является фактором, провоцирующим
паления, где ликвидируют внедрившиеся патоге-
развитие ряда заболеваний [1, 5].
ны и инициируют и поддерживают воспалитель-
Полифенолы относят к эффективным антиок-
ный процесс. Для этих целей нейтрофилы ис-
сидантам, способным ингибировать образование
пользуют различные эффекторные функции. В
АФК нейтрофилами как снаружи, так и внутри
частности, нейтрофилы фагоцитируют патогены
клетки [6, 7]. Тем не менее, мало что известно об
и клеточный дебрис, продуцируют активные
их способности влиять на другие функции ней-
формы кислорода (АФК), как снаружи клетки,
трофилов, такие как фагоцитоз и НЕТоз. Ранее
так и внутри фагосом, высвобождают, так назы-
высказывалось предположение, согласно которо-
ваемые внеклеточные ловушки, или NET (neutro-
му соединения, способные ингибировать АФК,
phil extracellular traps) [1,2]. Хотя данные функции
способны ингибировать и образование NET, за-
нейтрофилов важны для эффективной защиты
висимое от АФК [8]. В пользу данного предполо-
организма от патогенов, пролонгированная акти-
жения свидетельствуют некоторые данные, пред-
вация данных клеток играет важную роль в пато-
ставленные в литературе [8-10]. В частности, бы-
генезе ряда воспалительных и аутоиммунных за-
ло показано, что эпикатехин, катехин, рутин,
болеваний [3, 4]. Например, известно, что избы-
ресвератрол и кверцетин, проявляющие высокую
антиоксидантую активность, способны ингиби-
Сокращения: АФК - активные формы кислорода, NET - ровать АФК-зависимое образование NET, инду-
внеклеточные ловушки нейтрофилов (neutrophil extracellu-
цированное форбол-12-миристат-13-ацетатом
lar traps), DfTf - конъюгат таксифолина с глиоксалевой
кислотой, кальцеин-AM - кальцеин-ацетоксиметиловый
[8-10]. С другой стороны, образование NET, вы-
эфир.
званное отдельными стимулами, может быть не-
941
942
ШУБИНА и др.
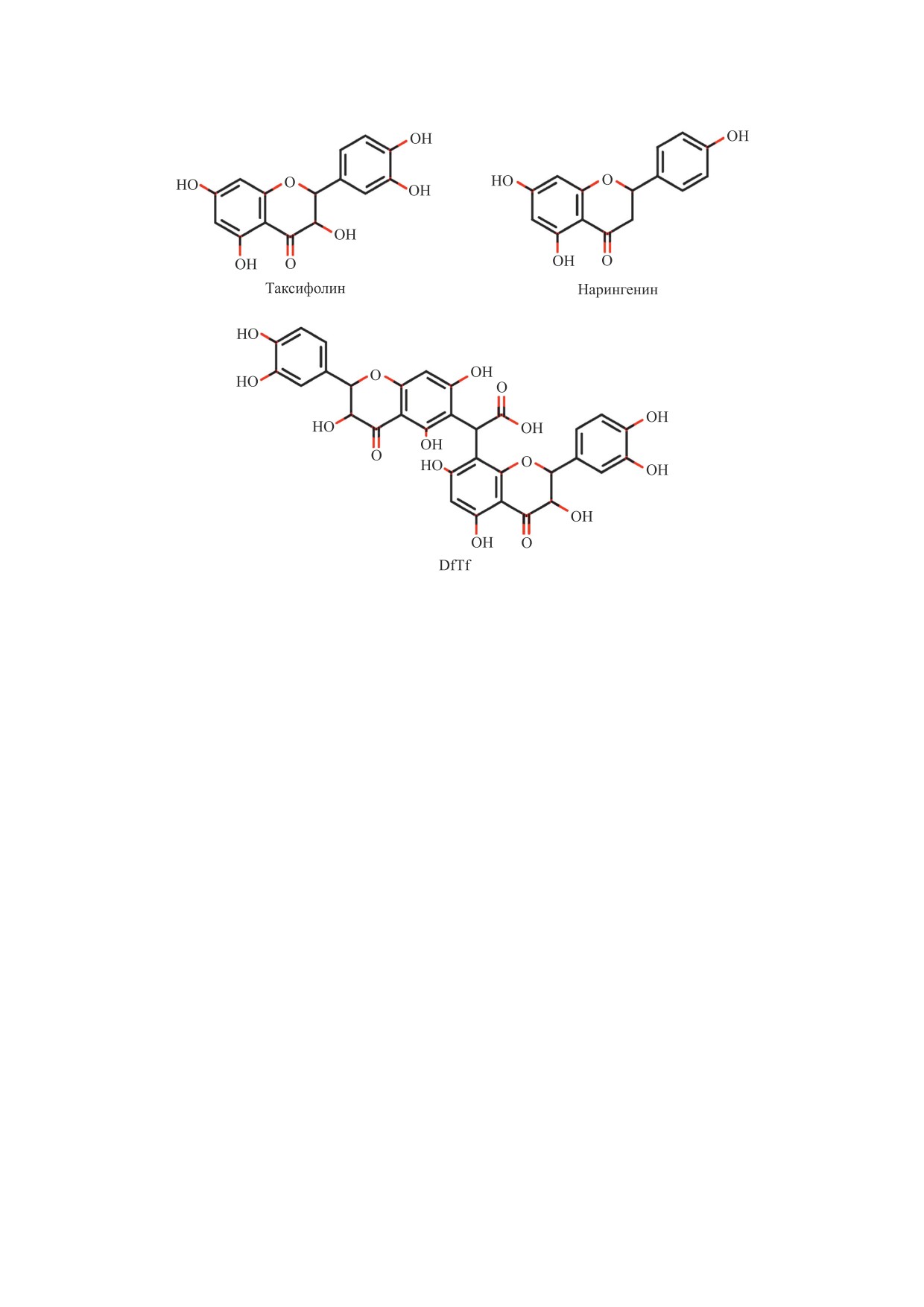
Рис. 1. Структуры исследуемых полифенолов. Конъюгат таксифолина с глиоксалевой кислотой обозначен как DfTf.
зависимым от АФК [11]. Так, образование NET,
фагоцитарную активность и адгезивные свойства
индуцированное латексными шариками, опсо-
нейтрофилов (рис. 1). Таксифолин и нарингенин
низированными IgG, происходило в присутствии
представляют собой природные полифенолы,
дифенилениодоний хлорида, сильного ингибито-
присутствующие в продуктах питания и напит-
ра NADPH-оксидазы, полностью останавливаю-
ках, среди которых цитрусовые и их соки, лике-
щего НЕТоз, индуцированный форбол-12-мири-
ры, вино и др. [16-18]. Данные соединения про-
стат-13-ацетатом [11]. Таким образом, вопрос о
являют разнообразные биологические свойства,
том каково влияние полифенолов на АФК-неза-
включая антиоксидантные, противовоспалитель-
висимый НЕТоз, остается открытым. Еще мень-
ные и противомикробные свойства [19-22]. DfTf
ше известно о влиянии полифенолов на фагоци-
является продуктом конденсации таксифолина и
тарную активность нейтрофилов. Отдельные дан-
глиоксалевой кислоты и, предположительно,
ные, встречающиеся в литературе, получены на
формируется в процессе хранения, обработки и
клеточных линиях и, в целом, свидетельствуют в
переваривания продуктов питания и напитков,
пользу того, что полифенолы могут оказывать как
содержащих данные соединения. О формирова-
усиливающее, так и ингибирующее действие на
нии продуктов конденсации флавоноидов с реак-
фагоцитарную активность клеток, которое, по
тивными карбонильными соединениями в про-
всей видимости, зависит от структуры данных со-
дуктах питания и in vivo уже сообщалось в литера-
единений [12, 13]. Другим открытым вопросом
туре [23, 24]. Глиоксалевая кислота формируется
остается влияние полифенолов на адгезивные
в вине, где, как известно, она реагирует с катехи-
свойства нейтрофилов. Как известно, благодаря
нами, приводя к образованию пигментов катиона
адгезивным свойствам возможна миграция ней-
ксантилия (xanthylium cation pigments). Довольно
трофилов из кровеносного русла к очагу воспале-
вероятно, что DfTf наряду с другими продуктами
ния и их надлежащее функционирование [14]. С
конденсации таксифолина также формируется в
другой стороны, чрезмерная адгезия нейтрофи-
процессе выдержки и хранения вина. Снижение
лов может приводить к повреждению собствен-
уровня таксифолина в вине со временем [18] так-
ных клеток и тканей [15].
же свидетельствует в пользу данного предполо-
жения.
Целью настоящей работы являлось изучение
влияния таксифолина, конъюгата таксифолина с
Хотя реакции полифенолов с реактивными
глиоксалевой кислотой (DfTf) и нарингенина на карбонильными соединениями протекают в про-
БИОФИЗИКА том 68
№ 5
2023
ВЛИЯНИЕ ТАКСИФОЛИНА, КОНЪЮГАТА ТАКСИФОЛИНА
943
цессе хранения, старения, обработки и перевари-
чего трижды отмывали раствором Хенкса. Иссле-
вания продуктов питания и напитков, богатых
дование проводили в 96-луночных планшетах
полифенолами, свойства таких продуктов и влия-
(SPL 30096, Южная Корея) следующим образом.
ние данных реакций на свойства полифенольной
В лунки, содержащие 100 мкл суспензии нейтро-
фракции остаются мало изученными. Действие
филов в растворе Хенкса (500 тыс. кл./мл) и эм-
образующихся продуктов на клетки также остает-
бриональную бычью сыворотку (10%), добавляли
ся в значительной степени неизвестными. В свя-
полифенолы, конечная концентрация которых
зи с вышесказанным исследование влияния DfTf
составляла 100 мкМ. Затем планшеты помещали в
на функциональную активность нейтрофилов
CO2-инкубатор на 30 мин. После этого к нейтро-
представляет собой отдельный интерес.
филам добавляли латексные частицы и инкуби-
ровали при 37°С в CO2-инкубаторе в течение двух
часов. За 30 мин до окончания инкубации клеток
МАТЕРИАЛЫ И МЕТОДЫ
с латексными шариками в среду добавляли ядер-
Синтез конъюгата таксифолина с глиоксалевой
ный краситель Hoechst
33342
(14533, Sigma,
кислотой. Синтез осуществляли по методике,
США) в концентрации 5 мкг/мл. В отдельных
описанной ранее [25]. Кратко: 148 мг глиоксале-
экспериментах, цитоплазму клеток окрашивали
вой кислоты и 20 мкл концентрированной соля-
кальцеин-ацетоксиметиоловым эфиром (кальце-
ной кислоты были добавлены к 608 мг таксифо-
ин-AM) (206700, Sigma, США). После этого клет-
лина, растворенного в 5 мл тетрагидрофурана.
ки однократно отмывали раствором Хенкса и фо-
Смесь нагревали при постоянном перемешива-
тографировали с помощью флуоресцентной мно-
нии с обратным холодильником в течение 2 ч.
гопараметрической флуоресцентной станции
Продукт конденсации выделяли в виде натриевой
ImageXpress MicroXL (Molecular Devices, США).
соли добавлением к смеси раствора гидроксида
Полученные снимки обрабатывали с использова-
натрия в этаноле. Продукт отмывали холодным
нием встроенного программного обеспечения.
спиртом и высушивали под вакуумом.
Долю клеток, вступивших в фагоцитоз, определя-
ли как отношение числа клеток, вступивших в
Выделение нейтрофилов. Нейтрофилы выделя-
фагоцитоз, к общему числу клеток. Среднее чис-
ли из цельной крови крыс-самцов линии Wistar
ло частиц, поглощенных клеткой, определяли как
методом лизиса эритроцитов с последующим
отношение общего числа поглощенных частиц к
центрифугированием на градиенте плотности
числу клеток, вступивших в фагоцитоз.
фикол-верографин (1.077/1.119) [26, 27]. Кратко:
к гепаринизированной крови добавляли двукрат-
Оценка адгезивных свойств нейтрофилов. По-
ный объем холодной (4°C) дистиллированной во-
сле съемки клеток с помощью флуоресцентной
ды и перемешивали в течении 20 с. После этого
многопараметрической флуоресцентной станции
быстро восстанавливали осмотичность раствора
ImageXpress MicroXL клетки инкубировали с
добавлением двукратного раствора фосфатно-со-
0.25%-м раствором трипсин-ЭДТА в течение
левого буфера. Далее суспензию клеток центри-
20 мин. Затем клетки двукратно отмывали рас-
фугировали при 180 g в течение 5 мин при 4°C.
твором Хенкса и оценивали общее число клеток,
Осадок отмывали раствором фосфатно-солевого
присутствующих в лунках, с помощью 96-луноч-
буфера. Полученную фракцию, обогащенную
ного микропланшетного ридера Infinite F200 (Te-
лейкоцитами, наслаивали на градиент фикол-ве-
can, Австрия). Измерения проводили при λex =
рографин (1.077/1.119) и центрифугировали в те-
= 360 нм, λem = 465 нм, что позволяет регистриро-
чение 15 мин при 180 g при 4°C. Фракцию, распо-
вать флуоресценцию, соответствующую ядерно-
ложенную на границе раздела фикол-верогра-
му красителю Hoechst 33342. За 100% принимали
фин (1.077/1.119), собирали и дважды отмывали
флуоресценцию нейтрофилов, не подвергавших-
раствором фосфатно-солевого буфера. Далее
ся каким-либо воздействиям. Количество адгези-
клетки ресуспендировали в растворе Хенкса (pH
рованных нейтрофилов выражали в процентах от
7.2) (106 клеток/мл). Содержание нейтрофилов во
исходного числа клеток.
фракции составляло не менее 96%.
Статистический анализ. Все эксперименты бы-
Оценка фагоцитарной активности. Фагоцитар-
ли выполнены трижды, в трех повторностях. Дан-
ную активность нейтрофилов оценивали методом
ные представлены как среднее значение ± стан-
флуоресцентной микроскопии. Для этих целей
дартное отклонение. Сравнение средних значе-
использовали латексные частицы, обладающие
ний проводили с помощью однофакторного
желто-зеленой флуоресценцией (L4530, Sigma,
дисперсионного анализа (ANOVA) с последую-
США), и латексные частицы, обладающие крас-
щим попарным сравнением с помощью Тьюки
ной флуоресценцией (L3030, Sigma, США). Сред-
теста. Для расчетов применяли статистический
ний размер частиц - 2 мкм. Частицы предвари-
программный пакет GraphPad Prism 9.0.0 (Graph-
тельно опсонизировали иммуноглобулином IgG
Pad Software Inc., США). Различия считали зна-
(I8015, Sigma, США) в течение одного часа, после
чимыми при p < 0.05.
БИОФИЗИКА том 68
№ 5
2023
944
ШУБИНА и др.
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
[30], предполагая вовлечение эндоплазматиче-
ского ретикулума в данный процесс. Таким обра-
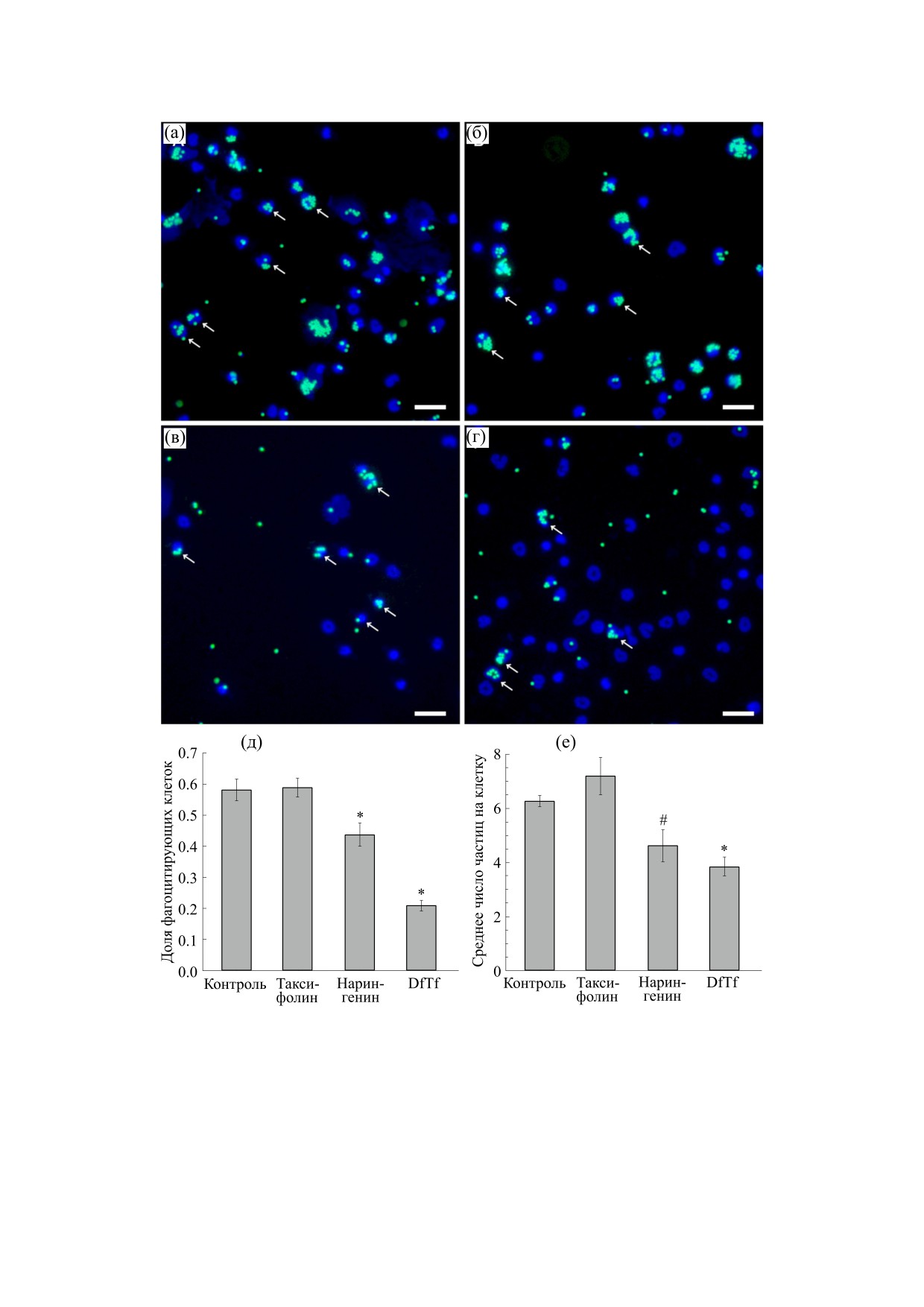
Для оценки фагоцитарной активности нейтро-
зом, природа окрашиваемых кальцеином-AM
филы инкубировали с латексными частицами
структур, а также механизмы формирования NET
L4530 (желто-зеленая флуоресценция), опсони-
остаются неясными на сегодняшний день и тре-
зированными IgG, в отсутствие (контроль) и в
буют дальнейшего изучения.
присутствии полифенолов (100 мкМ) в течение
На рис. 3 приведены флуоресцентные микро-
двух часов. Было показано, что таксифолин до-
фотографии нейтрофилов спустя два часа инку-
стоверно не влияет на фагоцитарную активность
бации с латексными частицами L3030 (красная
нейтрофилов, тогда как конъюгат DfTf значи-
флуоресценция), опсонизированными IgG, в от-
тельно снижает ее (рис. 2). В присутствии DfTf
сутствие (контроль) и в присутствии полифено-
снижается доля клеток, вступивших в фагоцитоз,
лов (100 мкМ). Видно, что во всех образцах на-
и среднее число частиц, поглощенных клеткой,
блюдается формирование структур, напоминаю-
на 64% и 39% соответственно (рис. 2д,е). Уста-
щих сеть. Наиболее выраженное образование
новлено, что другой исследуемый флавоноид, на-
данных структур наблюдается в контроле и в при-
рингенин, также снижает фагоцитарную актив-
сутствии нарингенина. Для сравнения на рис. 3
ность нейтрофилов, однако в меньшей степени,
также приведены данные о влиянии исследуемых
чем DfTf. В присутствии нарингенина доля кле-
полифенолов на фагоцитоз латексных частиц
ток, вступивших в фагоцитоз, снижалась прибли-
L3030 (красная флуоресценция) нейтрофилами.
зительно на 25%, как и среднее число частиц, по-
глощенных клеткой (рис. 2д,е).
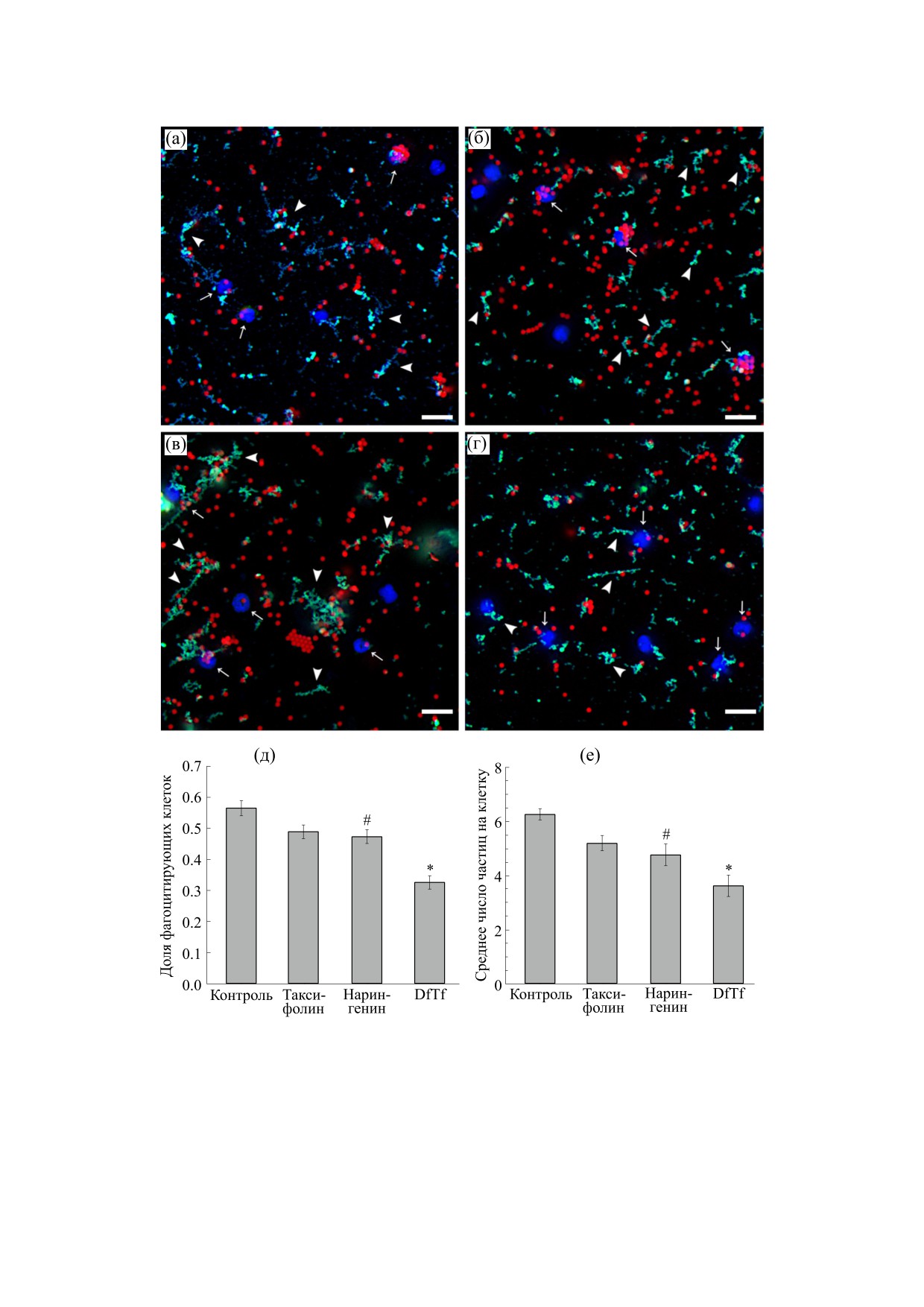
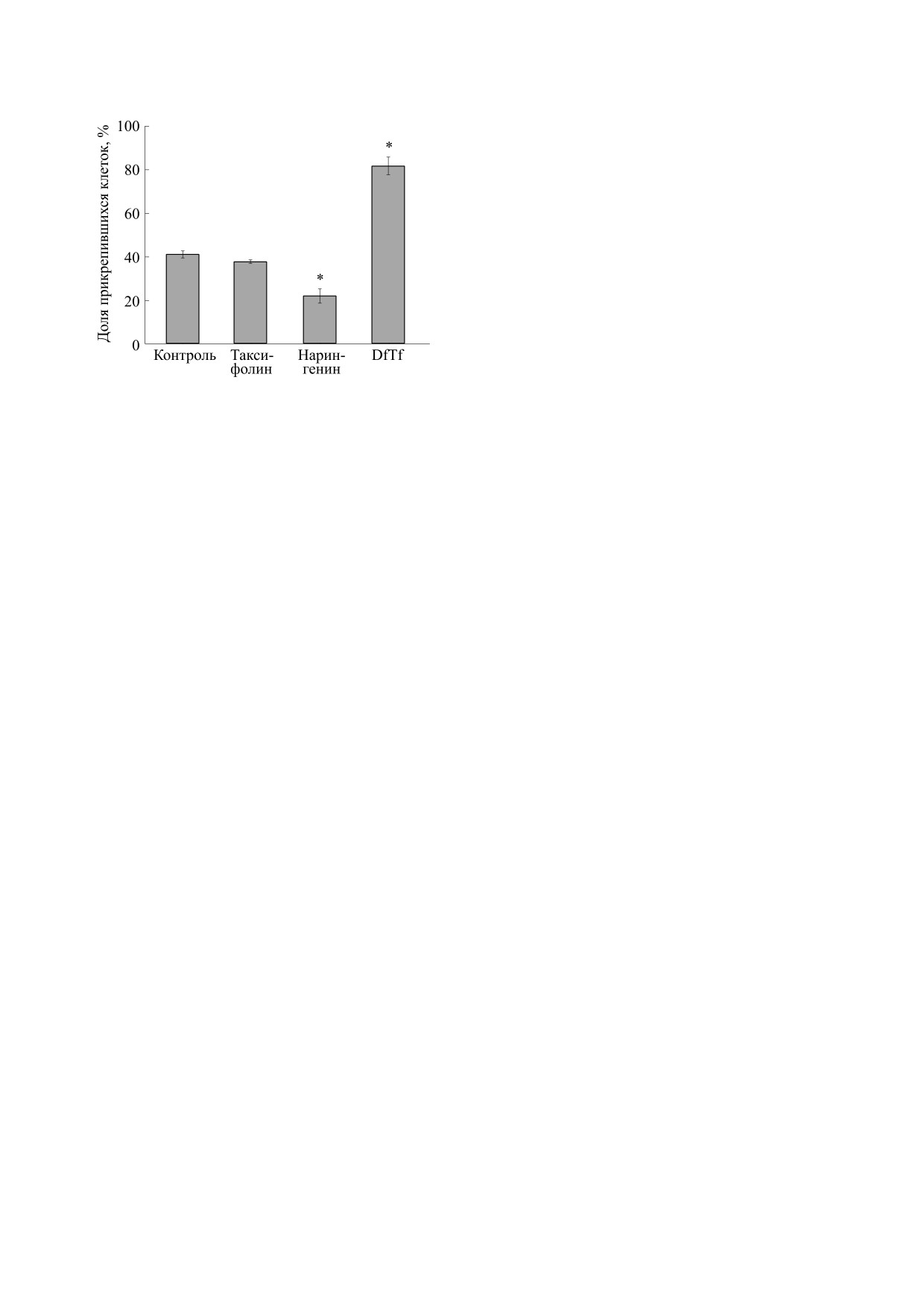
В рамках данной работы также было установ-
лено, что исследуемые полифенолы по разному
Важно отметить, что в процессе инкубации
влияют на адгезивные свойства нейтрофилов
клеток с латексными шариками, опсонизирован-
(рис. 4). Так, DfTf, несмотря на его ингибирую-
ными IgG, наблюдалось формирование структур,
щее влияние на фагоцитарную активность ней-
напоминающих сеть. Данные структуры окраши-
трофилов, усиливал адгезию данных клеток к по-
вались ядерным красителем Hoechst 33342 и,
верхности пластика, тогда как нарингенин инги-
предположительно, представляли собой NET
бировал ее. Влияния таксифолина на адгезивные
(рис. 3). Как уже упоминалось, о формировании
свойства нейтрофилов в условиях эксперимента
NET нейтрофилами человека в присутствии ла-
выявлено не было (рис. 4).
тексных шариков, опсонизированных IgG и ком-
плементом, ранее сообщалось в литературе [11].
Таким образом, полученные данные ука-
Интересно, что добавление в среду кальцеина-
зывают на то, что таксифолин, способный эф-
AM приводило к дополнительному окрашиванию
фективно ингибировать продукцию АФК ней-
данных структур, что также хорошо согласуется с
трофилами в ответ на различные стимулы [6, 7],
литературными данными, согласно которым
не оказывает достоверного влияния на фагоци-
NET окрашиваются таким красителем как Fluo-3
тарную активность и адгезию данных клеток. В то
AM [28]. Как известно, оба эти красителя прояв-
же время его структурный аналог нарингенин,
ляют флуоресценцию в присутствии ионов каль-
проявляющий меньшую антиоксидантную ак-
ция после отщепления эфирной группы внутри-
тивность [6, 7], оказывает ингибирующее дей-
клеточными эстеразами. Окрашивание формиру-
ствие на фагоцитоз и адгезию нейтрофилов. Это
ющейся сети данными красителями, вероятно,
позволяет говорить о том, что влияние данных
говорит о том, что кальций-содержащие структу-
структурно близких соединений на различные
ры входят в ее состав или солокализуются с ней.
функции нейтрофилов существенно отличается.
Примечательно, что митохондрии, являющиеся
При этом наблюдаемые ингибиторные эффекты
внутриклеточным кальциевым депо, также во-
указывают на противовоспалительный потенци-
влечены в НЕТоз. Митохондриальная ДНК была
ал обоих соединений. DfTf ингибирует фагоци-
обнаружена ассоциированной с ядерной ДНК в
тоз, но усиливает адгезию нейтрофилов к пласти-
составе NET [29, 30]. Однако механизмы, лежа-
ку в присутствии латексных частиц, опсонизиро-
щие в основе такой ассоциации, остаются невы-
ванных IgG, что свидетельствует о модуляции
ясненными [29, 30]. Можно предположить, что
функционального ответа нейтрофилов на внед-
митохондрии могут быть вовлечены в формиро-
рившийся патоген. Как известно, адгезивные
вание структур, окрашиваемых вышеупомянуты-
свойства нейтрофилов играют ключевую роль в
ми красителями. С другой стороны, нельзя ис-
функционировании данных клеток [14]. В част-
ключать вклад в образование данных структур
ности, адгезия нейтрофилов сопряжена с секре-
других органелл или компонентов клетки. На-
цией различных цитокинов, способствующих ин-
пример, обширная вакуолизация эндоплазмати-
фильтрации ткани макрофагами, поддержанию
ческого ретикулума, другого депо кальция в клет-
воспалительного процесса [31, 32], а также акти-
ке, наблюдалась на ранних стадиях НЕТоза, вы-
вации фибробластоподобных клеток, участвую-
званного иономицином и липополисахаридом
щих в процессе регенерации ткани [33]. С другой
БИОФИЗИКА том 68
№ 5
2023
ВЛИЯНИЕ ТАКСИФОЛИНА, КОНЪЮГАТА ТАКСИФОЛИНА
945
Рис. 2. Влияние исследуемых полифенолов на фагоцитоз латексных частиц L4530 (желто-зеленая флуоресценция)
нейтрофилами. (а-г) Репрезентативные флуоресцентные микрофотографии нейтрофилов в контроле (а), а также в
присутствии таксифолина (б), конъюгата таксифолина с глиоксалевой кислотой (в) и нарингенина (г). Стрелками
указаны отдельные клетки, вступившие в фагоцитоз. Масштабный отрезок 25 мкм. (д-е) Показатели фагоцитоза в
контроле и в присутствии исследуемых полифенолов: доля клеток, вступивших в фагоцитоз (д), среднее число частиц,
поглощенных клеткой (е). * Достоверные различия по сравнению с контролем, p < 0.001; # Достоверные различия по
сравнению с контролем, p < 0.05.
БИОФИЗИКА том 68
№ 5
2023
946
ШУБИНА и др.
Рис.
3. Влияние исследуемых полифенолов на фагоцитоз латексных частиц L3030 (красная флуоресценция)
нейтрофилами. (а-г) Репрезентативные флуоресцентные микрофотографии нейтрофилов в контроле (а), а также в
присутствии таксифолина (б), конъюгата таксифолина с глиоксалевой кислотой (в), и нарингенина (г). Стрелками указаны
отдельные клетки, вступившие в фагоцитоз. Головками стрелок указаны структуры, окрашиваемые Hoechst 33342 и
кальцеином-AM. Масштабный отрезок 25 мкм. (д-е) Показатели фагоцитоза в контроле и в присутствии исследуемых
полифенолов: доля клеток, вступивших в фагоцитоз (д), среднее число частиц, поглощенных клеткой (е). * Достоверные
различия по сравнению с контролем, p < 0.001; # Достоверные различия по сравнению с контролем, p < 0.005.
БИОФИЗИКА том 68
№ 5
2023
ВЛИЯНИЕ ТАКСИФОЛИНА, КОНЪЮГАТА ТАКСИФОЛИНА
947
ФИНАНСИРОВАНИЕ РАБОТЫ
Исследование выполнено при финансовой
поддержке Российского научного фонда (проект
№ 23-25-00149).
СОБЛЮДЕНИЕ ЭТИЧЕСКИХ СТАНДАРТОВ
Все манипуляции с подопытными животными
проводились в соответствии с международными
правилами обращения с лабораторными живот-
ными (Директива 2010/63/EU) и были одобрены
Комитетом по этике ИТЭБ РАН.
СПИСОК ЛИТЕРАТУРЫ
1.
Н. В. Воробьева, Вестн. Моск. ун-та. Сер. 16. Био-
Рис. 4. Влияние исследуемых полифенолов на адгезив-
логия, 75 (4), 210 (2020).
ные свойства нейтрофилов. * Достоверные различия по
2.
М. А. Челомбитько, Вестн. Моск. ун-та. Сер. 16.
сравнению с контролем, p < 0.001.
Биология, 73 (4), 242 (2018).
3.
М. Laforge, C. Elbim, C. Frère, et al., Nat. Rev. Im-
munol. 20 (9), 515 (2020).
стороны, нейтрофилы, вносят свой вклад и в за-
4.
A. Herrero-Cervera, O. Soehnlein, E. Kenne, Cell.
вершение воспаления, при этом их способность к
Mol. Immunol., 19 (2), 177 (2022).
адгезии также играет важную роль [32]. Прини-
5.
V. Papayannopoulos, Nat. Rev. Immunol., 18 (2), 134
мая во внимание вышесказанное, мы полагаем,
(2018).
что DfTf может представлять интерес в качестве
6.
D. Ribeiro, M. Freitas, S. M. Tomé, et al., Eur. J. Med.
компонента новых ранозаживляющих материа-
Chem., 67, 280 (2013).
лов.
7.
D. Ribeiro, E. Fernandes, and M. Freitas, In Flavo-
noids as Modulators of Neutrophils' Oxidative Burst:
Важно также отметить следующее. В рамках
Structure-Activity Relationship. Polyphenols: Mecha-
данной работы мы впервые показали, что конъ-
nisms of Action in Human Health and Disease (Acad.
югат таксифолина с глиоксалевой кислотой и ис-
Press, 2018), Chapter 20.
ходный флавоноид, таксифолин, по разному вли-
8.
T. Kirchner, E. Hermann, S. Möller, et al., Mediators
яют на фагоцитоз и адгезию нейтрофилов. Ранее
Inflamm., 2013, 710239 (2013).
мы обнаружили, что DfTf проявляет большую
9.
M. M. de Souza Andrade, V. N. C. Leal, I. G. Fer-
улавливающую активность по отношению к пе-
nandes, et al., Antioxidants (Basel), 11 (9),
1690
(2022).
роксиду водорода, чем таксифолин [25, 34]. В то
10.
G. S. Pereira, I. Percebom, S. Mendes, et al., Braz. J.
же время DfTf и таксифолин оказывали сопоста-
Biol., 84, e252936 (2022).
вимое антиоксидантное действие в бифазной си-
11.
M. Saffarzadeh, H. A. Cabrera-Fuentes, F. Veit, et al.,
стеме [34]. Таким образом, полученные результа-
Discoveries (Craiova), 2 (2), e19 (2014).
ты свидетельствуют о том, что свойства и биоло-
12.
M. Monobe, K. Ema, Y. Tokuda, et al., Cytotechnolo-
гическая активность полифенольной фракции
gy, 62 (3), 201 (2010).
могут существенно меняться, когда протекают
13.
S. Cui, J. Qian, P. Bo, J. Tradit. Chin. Med., 33 (6), 804
реакции конденсации полифенолов с реактивны-
(2013).
ми карбонильными соединениями.
14.
G. Berton, S. R. Yan, L. Fumagalli, et al., Int. J. Clin.
Lab. Res., 26 (3), 160 (1996).
В целом полученные данные указывают на то,
15.
А.А. Галкин и В.С. Демидова, Раны и раневые ин-
что полифенолы способны модулировать функ-
фекции. Журнал имени проф. Б.М. Костючёнка, 2
циональную активность нейтрофилов, что может
(2), 25 (2015).
играть важную роль в разрешении воспалитель-
16.
R. Cannataro, A. Fazio, C. La Torre, et al., Antioxi-
ного процесса, в частности когда пролонгирован-
dants (Basel), 10 (2), 328 (2021).
ная активация нейтрофилов приводит к повре-
17.
A. Rødtjer, L. Skibsted, and M. L. Andersen. Eur. Food
ждению клеток и тканей собственного организма.
Res. Technol., 223, 663 (2006).
18.
B.F. de Simón, M. Sanz, E Cadahía, et al., Food
Chem., 143, 66 (2014).
БЛАГОДАРНОСТИ
19.
J. Cai, H. Wen, H. Zhou, et al., Biomed. Pharmacoth-
er., 164, 114990 (2023).
Работа выполнена с использованием прибор-
20. Y. Liu, X. Shi, Y. Tian, et al., Front. Pharmacol., 14,
ной базы ЦКП ИТЭБ РАН. Авторы благодарны
1173855 (2023).
В.Г. Заикину (ЗАО “НПФ “Флавит”) за любезно
21. A. Duda-Madej, J. Stecko, J. Sobieraj, et al., Antibiot-
предоставленный таксифолин.
ics (Basel), 11 (11), 1628 (2022).
БИОФИЗИКА том 68
№ 5
2023
948
ШУБИНА и др.
22. D.Yang, R. Zhu, H.-X. Xu, et al., Food Bioscience, 53,
28. A. K. Gupta, S. Giaglis, P. Hasler, et al., PLoS One, 9
102811 (2023).
(5), e97088 (2014).
23. M. Chen, H. Zhou , C. Huang, et al., Food Chem.,
29. S. Yousefi, D. Stojkov, N. Germic, et al., Eur. J. Immu-
377, 132008 (2022).
nol., 49 (2), 221 (2019).
30. H. R. Thiam, S. L. Wong, D. D. Wagner, et al., Annu.
24. M. Chen, P. Liu, H. Zhou, et al., Front. Nutr., 9,
Rev. Cell. Dev. Biol., 36, 191 (2020).
973048 (2022).
31. A. Herrero-Cervera, O. Soehnlein, and E. Kenne, Cell.
25. V. S. Shubina and Yu. V. Shatalin, J. Food Sci. Tech-
Mol. Immunol., 19 (2), 177 (2022).
nol., 54 (6), 1467 (2017).
32. Baiula, R. Greco, L. Ferrazzano, et al., PLoS One, 15
26. M.T. Quinn and F.R. DeLeo, Neutrophil: Methods and
(8), e0237746 (2020).
Protocols. Methods in Molecular Biology, 2087 (Spring-
33. S. de Oliveira, E. Rosowski, and A. Huttenlocher, Nat.
er, 2020).
Rev. Immunol., 16 (6), 378 (2016).
27. N. Beloborodova, I. Bairamov, A. Olenin, et al., J.
34. V. S. Shubina, V. I. Kozina, and Yu. V. Shatalin, Anti-
Biomed. Sci., 19 (1), 89 (2012).
oxidants (Basel), 10 (8), 1262 (2021).
Effect of Taxifolin, a Conjugate of Taxifolin with Glyoxylic Acid, and Naringenin
on the Functional Activity of Neutrophils
V.S. Shubina, M.I. Kobyakova, and Yu.V. Shatalin
Institute of Theoretical and Experimental Biophysics, Russian Academy of Sciences,
Institutskaya ul. 3, Pushchino, Moscow Region, 142290 Russia
The aim of the present work was to study the effects of taxifolin, a conjugate of taxifolin with glyoxylic acid,
and naringenin on phagocytosis of latex beads by neutrophils and adhesive properties of these cells. It was
shown that taxifolin had no effect on the phagocytic activity and adhesive properties of neutrophils. A conju-
gate of taxifolin with glyoxylic acid strongly inhibited the phagocytic activity of neutrophils. At the same time,
the conjugate significantly enhanced adhesion of these cells. Naringenin decreased phagocytic activity of
neutrophils, though to a lesser extent than the conjugate did. Naringenin also inhibited adhesion of neutro-
phils. Thus, the data collected during our study indicate that polyphenols may influence the functional ac-
tivity of neutrophils that can play an important role in the modulation of inflammation, especially when pro-
longed activation of neutrophils leads to tissue damage.
Keywords: phagocytosis, adhesion, neutrophils, polyphenols
БИОФИЗИКА том 68
№ 5
2023