БИОФИЗИКА, 2022, том 67, № 3, с. 596-599
БИОФИЗИКА CЛОЖНЫX CИCТЕМ
УДК 577.3
ПУРИНЕРГИЧЕСКИЕ МЕХАНИЗМЫ АДАПТАЦИИ ДИАФРАГМЫ
МЫШИ В УСЛОВИЯХ АЛЛЕРГИЧЕСКОЙ ПЕРЕСТРОЙКИ
© 2022 г. А.Е. Хайруллин, А.Ю. Теплов, С.Н. Гришин, А.У. Зиганшин
Казанский государственный медицинский университет, 420012, Казань, ул. Бутлерова, 49
E-mail: khajrulli@yandex.ru
Поступила в редакцию 11.01.2022 г.
После доработки 11.01.2022 г.
Принята к публикации 24.01.2022 г.
В экспериментах in vitro на полоске диафрагмальной мышцы мыши методом изометрической
электростимуляционной механомиографии выявлена роль синаптической модуляции в процессах
адаптации двигательной единицы к аллергии. Различия в сократительной активности мышцы под
влиянием экзогенной АТФ у интактных и сенсибилизированных яичным альбумином животных
демонстрируют участие пуринергических механизмов в развитии приспособительных изменений.
В процессе сенсибилизации взаимодействие АТФ-зависимых механизмов как пост-, так и
пресинаптической мембраны обеспечивает адекватную перестройку сократительной функции
дыхательных мышц к бронхоконстрикции.
Ключевые слова: m. diaphragma, мышь, АТФ, белковая сенсибилизация, аллергия.
DOI: 10.31857/S0006302922030206, EDN: APYFUH
Механизмы адаптации поперечнополосатых
влиянии экзогенной АТФ на мышцы интактных
мышц, обеспечивающих внешнее дыхание, при
и сенсибилизированных животных коррелируют
бронхиальной астме остаются до конца не изу-
с вектором изменения тока неквантовой секре-
ченными. Актуальность этой проблемы опреде-
ции ацетилхолина [3]. Пресинаптические струк-
ляется как широким распространением аллерги-
туры не могут оставаться непричастными к про-
ческих заболеваний, так и вопросами спортивной
цессу адаптации, однако их вовлеченность в
медицины. В частности, хорошо известно, что
описанных условиях остается совершенно неизу-
среди участников олимпийских сборных сканди-
ченной.
навских стран по зимним видам спорта встреча-
Цель данной работы заключалась в изучении
ется много астматиков [1, 2]. При обструкции
роли пуринергической синаптической модуля-
воздухоносных путей в условиях повышения
ции диафрагмы (m. diaphragma) мыши в адапта-
внешней нагрузки и развития вслед за этим гипо-
ции к аллергической перестройке.
ксии дыхательная мускулатура для компенсации
возникающей кислородной недостаточности вы-
МЕТОДЫ
нуждена работать интенсивнее. За счет чего про-
исходит эта интенсификация, до конца не ясно. В
Подготовительные процедуры. Эксперименты
предыдущих исследованиях нами было показано,
проводили на изолированных препаратах m. dia-
что адаптация дыхательных мышц в условиях
phragma 24 белых мышей обоего пола массой 25-
сенсибилизации обеспечивается АТФ-зависимы-
32 г. Животных содержали в специальных боксах
ми механизмами, реализуемыми на постсинапти-
на обычном пищевом режиме, со свободным до-
ческой мембране [3]. При белковой сенсибили-
ступом к воде. Мышей сенсибилизировали яич-
зации АТФ как комедиатор синаптической
ным альбумином [3]. В качестве контроля ис-
передачи нервно-мышечного соединения в зна-
пользовали несенсибилизированных животных.
чительной степени определяет динамику сокра-
Для эксперимента мыши были обескровлены под
тительной функции поперечнополосатых мышц.
анестезией (этаминал натрия, 40 мг/кг), после че-
В этих работах показана роль постсинаптическо-
го из центральной части левой m. diaphragma вы-
го звена реализации механизмов адаптации, на
деляли полоску шириной 2-3 мм и длиной 15-
котором определяется его чувствительность к хо-
18 мм.
линомиметику. В частности, различия в измене-
Условия проведения экспериментов по реги-
нии силы сокращения на карбахолин in vitro при страции параметров сокращения. Мышечный
596
ПУРИНЕРГИЧЕСКИЕ МЕХАНИЗМЫ АДАПТАЦИИ ДИАФРАГМЫ МЫШИ
597
ночку с раствором Кребса добавляли раствор
АТФ в концентрации 100 мкМ, через 10 мин ин-
кубации записывали сократительные ответы на
электрическую стимуляцию. Далее ткань промы-
вали несколько раз и инкубировали в течение
20 мин антагонистом (сурамином в концентра-
ции 100 мкМ). После этого добавляли агонист
(АТФ 100 мкМ) и повторно записывали сократи-
тельные ответы. Все сократительные ответы вы-
числяли в процентах от исходных сокращений.
Достоверность различий. Статистическую об-
работку данных проводили с помощью програм-
мы статистической обработки SPSS Statistics.
Проверку соответствия полученных данных нор-
мальному распределению проводили с помощью
критерия Колмогорова. Рассчитывали средние
арифметические анализируемых параметров и
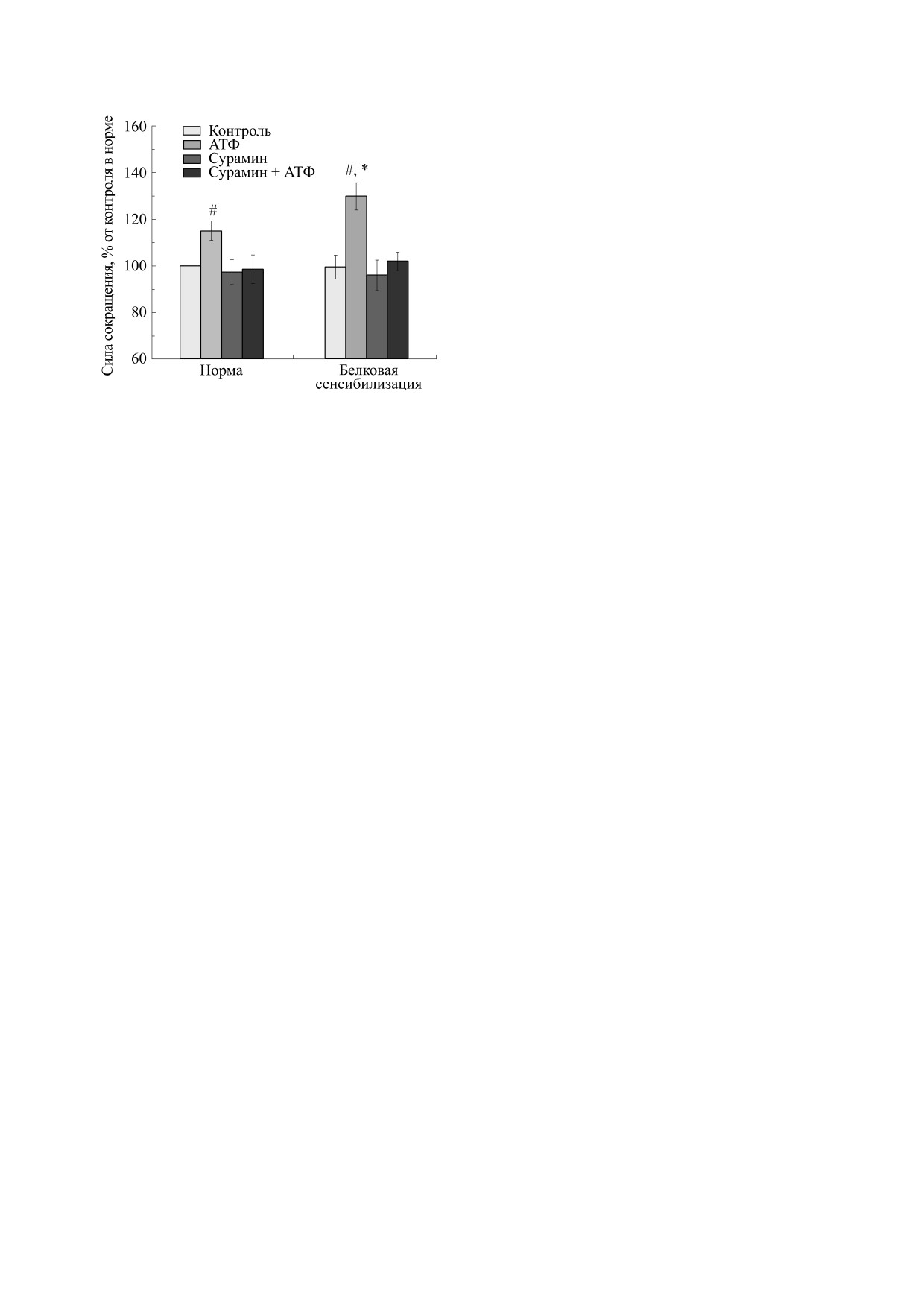
Сила сокращений m. diaphragma мыши, вызванных
стандартную ошибку. Статистическую значи-
электрической стимуляцией интактных
(«Норма»,
мость наблюдаемых изменений оценивали с по-
n=14) и сенсибилизированных («Белковая сенсибили-
мощью критерия Стьюдента для независимых и
зация», n = 10) животных в отсутствие и в присутствии
попарно сопряжeнных выборок. Различия рас-
АТФ (100 мкМ) и сурамина (100 мкМ). Результаты
сматривались как значимые при p < 0.05.
представлены в виде M ± m в % от исходных величин,
принятых за 100%; # - р < 0.05 от контроля в норме,
*- р< 0.05 от эффекта АТФ в норме.
РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЯ
Электростимуляция полоски m. diaphragma
препарат помещали вертикально в термостатиру-
сенсибилизированного животного практически
емую ванночку объемом 10 мл. Один конец мыш-
не вызывала изменения силы ее сокращения в
цы был зафиксирован, другой конец при помощи
сравнении с контролем. Если у последних она со-
лигатуры крепили к датчику механической актив-
ставляла 542 ± 24 мг (n = 14), то в случае белковой
ности. Ванночку заполняли модифицированным
сенсибилизации - 538 ± 27 мг (n = 10). Влияние
раствором Кребса следующего состава (в мМ):
экзогенного АТФ на мышцы обеих групп показа-
NaCl - 118.0, KCl - 4.75, CaCl2 - 2.5, NaHCO3 -
ло следующую динамику. У несенсибилизиро-
24.8, KH2PO4
-
1.18, MgSO4·7H2O
-
1.18,
ванных мышей сила сокращения повышалась до
глюкоза - 11.0, рН 7.4 ± 0.1. Условия изометрии
623 ± 23 мг, т.е. до 115.1 ± 4.2% (n = 14), а в усло-
виях белковой сенсибилизации - до 703.5 ±
достигались растяжением препарата мышцы в
± 31.4 мг, т.е. до 129.8 ± 5.8% (n = 10). Более выра-
ванночке в течение 20 мин с силой 0.5 г при по-
женное потенцирование сокращения дыхатель-
стоянной перфузии раствором при температуре
ной мышцы после влияния АТФ у второй группы
37°С. Сократительную реакцию изолированной
в сравнении с первой (115.1% против 129.8%;
мышцы вызывали при помощи электрического
p < 0.05) (см. рисунок) свидетельствует об участии
стимулятора MultiStim D330 (Digitimer, Велико-
пуринергических механизмов пресинаптических
британия) [2, 3]. Прямоугольные электрические
образований в адаптации m. diaphragma к белко-
импульсы частотой 0.1 Гц и длительностью 0.5 мс
вой сенсибилизации. Инкубация мышцы с сура-
наносили в течение двух минут (двенадцать пи-
мином (100 мкМ) полностью предупреждала вли-
ков). Среднее значение силы всех двенадцати со-
яние АТФ на силу сокращений.
кращений анализировали как один результат.
Силу сокращения оценивали в граммах. Ответы
записывали с помощью изометрического механи-
ОБСУЖДЕНИЕ
ческого датчика FSG-01 (Linton Instrumentation,
В представленной работе показана возмож-
Великобритания) и аналого-цифрового преобра-
ность комплексного подхода к изучению адапта-
зователя MP100WSW (Biopack, США) [4, 5]. Ре-
ционных механизмов, которые развиваются в ды-
зультаты исследования наблюдали на мониторе и
сохраняли на персональном компьютере.
хательной мышце в условиях экспериментальной
патологии. Дана дифференцированная оценка
Эффекты пуринергических агонистов и антаго-
степени участия АТФ-зависимых механизмов в
нистов. В начале эксперимента сократительные
различных отделах мионеврального соединения.
ответы диафрагмы на электрическую стимуля-
Использованная экспериментальная модель поз-
цию регистрировали дважды с интервалом 5 мин
воляет выявлять преимущественно пресинапти-
(для оценки стабилизации ответов). Затем в ван-
ческие эффекты пуринов, постсинаптические
БИОФИЗИКА том 67
№ 3
2022
598
ХАЙРУЛЛИН и др.
эффекты становятся более выражены в условиях
ющая импульсацию по аксону, не приводит к
гипотермии [4-7].
появлению гиперчувствительности к гистамину,
что делает невозможным ее сокращение на этот
Результаты исследований показали, что у диа-
медиатор. Действие трофических факторов и им-
фрагмальной мышцы мыши в ходе аллергической
пульсная активность мотонейрона регулирует
перестройки при электростимуляции в присут-
синтез белков гистаминовых рецепторов по неза-
ствии экзогенной АТФ механизмы реализации
висимым механизмам [10]. Вероятно, таким об-
сократительной функции претерпевают суще-
разом белковая сенсибилизация изменяет функ-
ственные изменения. Как известно, кроме вы-
циональные характеристики мионеврального пе-
полнения в организме роли макроэрга АТФ явля-
рехода за счет увеличения чувствительности
ется комедиатором синаптической передачи [8].
постсинаптической терминали к холиномимети-
В предыдущих работах нами было показано, что в
ку и гистамину. Кроме этого, в немногочислен-
этих условиях пуринергические механизмы
ных источниках говорится о частичном измене-
участвуют в изменении чувствительности постси-
нии гистохимического профиля белкового соста-
наптической мембраны диафрагмальной мышцы
ва мышц в результате белковой сенсибилизации,
к холиномиметику [9]. В настоящем исследова-
которое заключается в небольшом увеличении
нии продемонстрировано, что АТФ-зависимые
«быстрых» мышечных волокон [11, 12].
процессы, локализованные и в пресинаптиче-
ском образовании, также вносят свой вклад в ре-
Описанные механизмы определяют способ-
ализацию механизмов адаптации в условиях бел-
ность организма противостоять гипоксии при
ковой сенсибилизации.
ожидаемом снижении проходимости воздухонос-
Если сама белковая сенсибилизация не оказы-
ных путей. Очевидно, что при реализации функ-
вает значимого действия на силу сокращения
ции внешнего дыхания АТФ-зависимые меха-
диафрагмы, то инкубация с экзогенной АТФ при-
низмы в различных отделах мионеврального со-
водила к росту этого показателя, причем у живот-
единения взаимно дополняют друг друга. Этим в
ных сенсибилизированной и контрольной групп
условиях необходимого роста работоспособности
степень этого увеличения существенно различа-
обеспечивается адаптация сократительных струк-
лась. Обнаружено возрастание этого показателя у
тур диафрагмы. Таким образом, поддерживается
сенсибилизированных животных по сравнению с
адекватный режим внешнего дыхания в условиях
контрольными, что свидетельствует об участии
аллергической перестройки, что в свою очередь
пуринергических механизмов в процессах адап-
позволяет организму заранее подготовиться к
тации мышцы к условиям белковой сенсибили-
предстоящим изменениям.
зации.
Различие динамики показателей, характеризу-
ВЫВОДЫ
ющих пресинаптическое образование демон-
стрирует комплексную картину участия пуринов
Адаптация дыхательных мышц при аллергиче-
в адаптации мионеврального соединения к аллер-
ской перестройке обеспечивается пуринергиче-
гической перестройке организма.
скими механизмами и заключается в готовности
Диафрагмальная мышца во время подготовки
противостоять перспективной гипоксии, возни-
к перспективной бронхоконстрикции внешне
кающей вследствие бронхоспастического син-
практически не меняет своих сократительных
дрома. Рост работоспособности m. diaphragma
свойств. Однако начинают развиваться механиз-
мыши, компенсирующей кислородную недоста-
мы, которые в перспективе обеспечат повышение
точность при обструкции воздухоносных путей,
ее работоспособности. Участие АТФ как комеди-
обеспечивается АТФ-опосредованными процес-
атора в работе и пре-, и постсинаптической мем-
сами как на уровне пре-, так и постсинаптиче-
браны свидетельствует о сложных взаимоотно-
ских структур этой мышцы.
шениях между этими процессами в различных ча-
стях мионеврального соединения, что регулирует
сократительную деятельность дыхательной мыш-
КОНФЛИКТ ИНТЕРЕСОВ
цы при аллергических реакциях.
Авторы заявляют об отсутствии конфликта
Как известно, клетками-мишенями первого
интересов.
порядка в анафилактическом сокращении ске-
летных мышц теплокровных являются тучные
клетки как источник гистамина, необходимого
СОБЛЮДЕНИЕ ЭТИЧЕСКИХ СТАНДАРТОВ
для реализации анафилактического сокращения
мышц. Чувствительность скелетных мышц к ги-
Все применимые международные, националь-
стамину регулируется нейротрофическим кон-
ные и институциональные принципы ухода и ис-
тролем; блокада аксонального транспорта аппли-
пользования животных при выполнении работы
кацией колхицина на седалищный нерв, сохраня-
были соблюдены.
БИОФИЗИКА том 67
№ 3
2022
ПУРИНЕРГИЧЕСКИЕ МЕХАНИЗМЫ АДАПТАЦИИ ДИАФРАГМЫ МЫШИ
599
СПИСОК ЛИТЕРАТУРЫ
7. A. E. Khairullin, A. U. Ziganshin, and S. N. Grishin,
Biophysics 65, 858 (2020).
1. P. Kippelen, F. Friemel, and P. Godard, Rev. Mal. Re-
spir. 20 (3), 385 (2003).
8. A. U. Ziganshin, A. E. Khairullin, C. H. V. Hoyle, and
2. L. P. Boulet and P. M. O’Byrne, New Engl. J. Med.
S. N. Grishin, Int. J. Mol. Sci. 21, 6423 (2020).
372 (7), 641 (2015).
9. А. Ю. Теплов, А. М. Фархутдинов, В. И. Торшин
3. A. Y. Teplov, S. N. Grishin, M. A. Mukhamedyarov,
и др., Вестн. новых мед. технологий 21, 6 (2014).
et al., Exp. Physiol. 94 (2), 264 (2009).
10. V. V. Valiullin and A. M. Devyataev, Allergy 50, 381
4. A. U. Ziganshin, A. E. Khairullin, V. V. Zobov, et al.,
(1995).
Muscle & Nerve 55 (3), 417 (2017).
11. I. M. Vikhlyantsev and Z. A. Podlubnaya, Biophysics
5. A. U. Ziganshin, A. E. Khairullin, A. Y. Teplov, et al.,
53 (6), 592 (2008).
Muscle & Nerve 59 (4), 509 (2019).
6. A. E. Khairullin, A. Yu. Teplov, S. N. Grishin, et al.,
12. A. M. Devyataev and V. V. Valiullin, Bull. Exp. Biol.
Biophysics 64, 812 (2019).
Med. 117 (2), 194 (1994).
Purinergic Mechanisms in the Adaptation of Mouse Diaphragm to Allergic Disorders
A.E. Khairullin, A.Yu. Teplov, S.N. Grishin, and A.U. Ziganshin
Kazan State Medical University, ul. Butlerova 49, Kazan, 420012 Russia
In vitro study of a muscle strip from mouse diaphragm, while performing isometric contractions using mech-
anomyography during the application of electrical stimulation, revealed that synaptic modulation contributes
to the processes of adaptation of the motor unit to allergy. Differences in muscle contractile activity under the
influence of exogenous ATP in intact and egg albumin-sensitized animals demonstrate the involvement of
purinergic mechanisms in the development of adaptive changes. In the process of sensitization, the interac-
tion between pre- and postsynaptic ATP-dependent mechanisms ensures proper rearrangements in the con-
tractile function of the respiratory muscles to bronchoconstriction.
Keywords: m. diaphragma, mouse, ATP, protein sensitization, allergy
БИОФИЗИКА том 67
№ 3
2022