БИОФИЗИКА, 2022, том 67, № 3, с. 540-548
БИОФИЗИКА КЛЕТКИ
УДК 57.05
СТОХАСТИЧНОСТЬ ЭКСПРЕССИИ БЕЛКА p53 В КЛЕТКАХ ПЕРВИЧНЫХ
И ПЕРЕВИВАЕМЫХ ЛИНИЙ ЧЕЛОВЕКА
© 2022 г. Р.А. Ковалёв, Н.Д. Федорова, Р.А. Пантина, Е.В. Семенова,
М.В. Филатов, Е.Ю. Варфоломеева
Петербургский институт ядерной физики им. Б.П. Константинова Национального исследовательского центра
«Курчатовский институт», Россия, 188300, Гатчина, Ленинградская обл., мкр. Орлова роща, 1
E-mail: varfolomeeva_ey@pnpi.nrcki.ru
Поступила в редакцию 24.03.2022 г.
После доработки 24.03.2022 г.
Принята к публикации 30.03.2022 г.
Стохастическая экспрессия генов может иметь важные последствия для судьбы как каждой кон-
кретной клетки, так и клеточной популяции в целом. В данной работе мы демонстрируем значи-
тельную вариабельность экспрессии p53, вплоть до полного ее отсутствия, в клетках первичных гли-
областом человека, чего не наблюдается в клетках нормальных фибробластов ФЛЭЧ и перевивае-
мых опухолевых линий, таких как HeLa и HT1080. При этом вариабельность экспрессии p53 в
клетках зависит от густоты посева. Показываем, что при подавлении транскрипции актиномици-
ном Д или нокдауне гена p53 РНК-интерференцией восстановление его уровня экспрессии в клет-
ках проходит стохастически.
Ключевые слова: стохастическая клеточная гетерогенность, экспрессионный шум, p53, первичные
глиобластомы.
DOI: 10.31857/S0006302922030127, EDN: AODLQO
чевую роль в определении судьбы клетки при по-
Экспрессия генов - это стохастический про-
вреждениях играет белок p53 [7]. Отсутствие бел-
цесс, в результате которого в популяции генети-
ка p53 в клетках затрудняет своевременный ответ
чески идентичных клеток могут возникнуть зна-
на клеточные стрессы, способствуя онкогенезу.
чительные межклеточные расхождения в уровнях
Очевидно, что анализ стохастических процессов
мРНК и белков. Неоднородность экспрессии мо-
при экспрессии гена р53 в опухолевых клетках
жет иметь важные последствия для судьбы как
представляет не только научный, но и большой
каждой конкретной клетки, так и клеточной по-
практический интерес, поскольку экспрессион-
пуляции в целом [1-4]. Клетки разнообразных
ный профиль может контролироваться различ-
организмов от микробов до высших постоянно
ными стратегиями регуляции генов, приводящим
подвергаются воздействию эндогенных и экзо-
к режимам с низким уровнем шума [8].
генных стрессов, приводящих к генетическим по-
вреждениям, эпигенетическим отклонениям и
Наиболее подходящей методологией исследо-
ошибкам регуляторных систем, что, в конечном
вания стохастической экспрессии генов является
итоге, и обуславливает стохастический характер
мониторинг продукции, деградации и функцио-
экспрессии генов [2, 5]. Экспрессионный шум
нального состояния биомолекул в реальном вре-
способствует формированию различных феноти-
мени в живых клетках [9]. В подавляющем боль-
пов и улучшает способность приспосабливаться к
шинстве работ уровень экспрессии p53 оценива-
изменяющимся условиям на уровне популяции,
ют, используя вестерн-блот либо полимеразную
являясь, по-видимому, важной составляющей
цепную реакцию, однако эти методы дают лишь
эволюционного прогресса. Однако в некоторых
усредненное значение уровня экспрессии, не от-
случаях стохастические флуктуации экспрессии
ражая поклеточного распределения. Конфокаль-
могут вызывать индукцию онкогенов или нару-
ная микроскопия и цитометрия же позволяют
шение регуляции генов-супрессоров опухолей,
выявить неоднородность экспрессионных про-
облегчая адаптивные программы злокачествен-
филей внутри популяции.
ной трансформации в условиях стресса [6]. Клю-
В этой работе мы изучали экспрессию гена p53
Сокращение: GFP - зеленый флуоресцентный белок (green
в клетках различных линий человека, как в пере-
f luorescent protein).
виваемых, так и в первичных. Продемонстриро-
540
СТОХАСТИЧНОСТЬ ЭКСПРЕССИИ БЕЛКА p53
541
вана стохастическая изменчивость экспрессии
центный белок GFP, проводили с помощью про-
гена p53 в популяции при внесении возмущений в
точного цитофлуориметра Cell Lab Quanta SC
функционирование клеток. Используя покадро-
фирмы Beckman Coulter (США). Сортировка кле-
вую микроскопию и проточную цитометрию, мы
ток по уровню флуоресценции GFP-белка осу-
исследовали кинетику такой изменчивости экс-
ществлялась с помощью сортера фирмы Beckman
прессии генов в клетках. Также нами показана
Coulter (США).
значительная вариабельность экспрессии p53 в
Анализ экспрессии генов в режиме реального
первичных клетках глиобластомы.
времени в живых клетках по технологии Smart-
Flare. Для прижизненного наблюдения за уров-
нем и динамикой включения экспрессии генов в
МАТЕРИАЛЫ И МЕТОДЫ
клетках (оценка концентрации РНК) использо-
Клеточные линии и условия культивирования.
вали набор SmartFlareтм (Merck Millipore, Герма-
Работа проведена на первичных и перевиваемых
ния). Согласно инструкции производителя зонды
культурах клеток человека: ФЛЭЧ (фибробласты
SmartFlareтм добавляли к исследуемому образцу,
легкого эмбриона человека), HT-1080 (фибросар-
спустя 4 ч инкубации с использованием конфо-
кома) и HeLa (аденокарцинома шейки матки) -
кального микроскопа (Leica, Германия) реги-
стандартные перевиваемые клеточные линии с
стрировали флуоресценцию в клетках.
диким типом белка p53 (Коллекция клеточных
культур Института цитологии РАН, Санкт-Пе-
Процедура подавления транскрипции РНК с по-
тербург), и Gl-La, Gl-Tr - первичные культуры
мощью актиномицина Д. Исследуемый образец
глиомных клеток, полученные в нашей лаборато-
клеток инкубировали в полной среде с добавле-
рии, а также на клеточной линии GFP-p53ΔY126-
нием актиномицина Д в концентрации 0.1 мкг/мл
HEK293, полученной нами путем введения плаз-
в течение 14 ч. Далее добавляли зонды Smart-
миды p53ΔY126-GFP в клетки линии HEK293 с
Flareтм на 4 ч и затем регистрировали флуорес-
последующим клонированием [10].
ценцию на конфокальном микроскопе. После
Клетки культивировали в полной среде
этого заменяли полную среду с актиномицином Д
(DMEM/F12 (ООО «Биолот», Санкт-Петербург)
на полную среду без него, снова добавляли зонды
с добавлением 10% эмбриональной сыворотки
SmartFlareтм и наблюдали за динамикой флуорес-
крупного рогатого скота (Biowest, Франция)), без
ценции образца в течение суток (для регистрации
антибиотиков, в 5%-й СО2-атмосфере при 37°С.
флуоресценции использовался конфокальный
микроскоп).
Для получения клонов (потомков одной клет-
ки) клетки линии GFP-p53ΔY126-HEK293 рассе-
Система РНК-интерференции для подавления
вали в 96-луночные планшеты. Один из наиболее
экспрессии гена ТР53. Подавление экспрессии ге-
ярких по уровню флуоресценции меченного зеле-
на p53 проводили с помощью трансфекции ко-
ным флуоресцентным белком (GFP - Green Flu-
ротких интерферирующих молекул РНК (ан-
orescent Protein) клон (далее в тексте GFP-клон) с
тиP53-siРНК) через 24 ч после посева клеток в
высоким уровнем экспрессии белка GFP-
чашки с оптическим дном. Трансфекцию ан-
p53ΔY126 был использован в дальнейшей работе.
тиP53-siРНК (ON-TARGETplus SMARTpool,
Ранее было показано, что в клетках GFP-клона
Dharmacon, США) в концентрации 100 нM в 1×
детектируются как белок GFP-p53ΔY126, так и
трансфекционном буфере (Dharmacon, США)
белок p53 дикого типа [11]. В дальнейшем пересев
проводили с использованием липофектамина
клеток осуществляли по мере образования клет-
(Dharmacon-FECT) по методике производителя
ками монослоя. Клетки обрабатывали раствором
Версен/трипсин 1:1 (ООО «Биолот», Санкт-Пе-
тербург) в течение 10 мин, переносили в центри-
фужную пробирку, осаждали
5 мин при
РЕЗУЛЬТАТЫ
1000 об/мин, ресуспендировали в питательной
Стохастическая экспрессия белка GFP-
среде и 1/20 часть суспензии клеток переносили в
p53ΔY126 в клетках линии GFP-p53ΔY126-
свежий флакон. Анализ полученных клеточных
HEK293. Плазмида p53ΔY126-GFP кодирует му-
линий (клонов) и отбор линий с высоким содер-
тантный белок р53 с делецией тирозина в 126-м
жанием GFP проводили на цитометре Cell
положении пептидной цепи белка, несущий на
LabQuanta (Beckman Coulter, США). Для прижиз-
N-конце зеленый флуоресцентный белок (GFP).
ненного наблюдения клетки сеяли в чашки диа-
Мутантная форма белка р53 обладает некоторы-
метром 35 мм с оптическим дном (MatTek Сorp.,
ми функциями белка p53 дикого типа. В частно-
США) и визуализировали на конфокальном мик-
сти, белок p53ΔY126 может работать как тран-
роскопе Leica SP5 (Leica, Германия).
скрипционный фактор, активируя индукцию
Проточная цитометрия и сортировка клеток. Ре-
белка p21, и вызывать клеточную гибель, однако
гистрацию количества клеток, несущих флуорес-
по сравнению с диким типом белка р53 его актив-
БИОФИЗИКА том 67
№ 3
2022
542
КОВАЛЁВ и др.
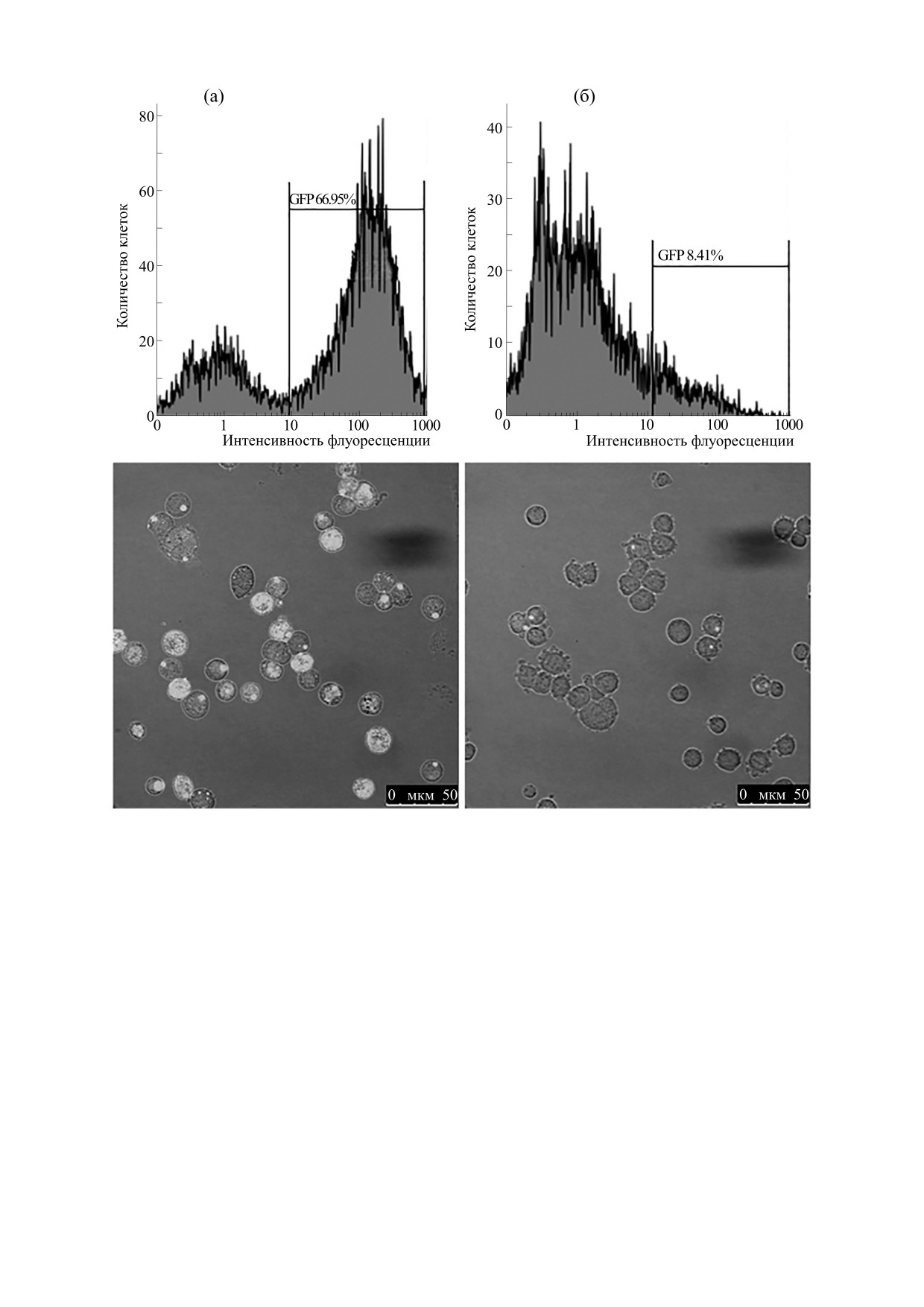
Рис. 1. Экспрессия белка GFP-p53ΔY126 в клетках различных клонов HEK293.
ность снижена [11]. Исследование клона линии
p53ΔY126 GFP от клетки к клетке в каждом клоне
HEK293, экспрессирующего белок GFP-
(рис. 1а,б).
p53ΔY126, показало существенное различие экс-
Чтобы показать, что в конкретной клетке кон-
прессии данного белка в различных клетках кло-
центрация исследуемого белка GFP-p53ΔY126
на от высокой до полностью отсутствующей. Что-
может меняться с течением времени, мы с помо-
бы исключить зависимость наблюдаемой нами
щью конфокального микроскопа в течение 72 ч
разницы в концентрации GFP-p53ΔY126 как от
вели наблюдение за живыми клетками линии
потери встроенной плазмиды во время роста кле-
GFP-p53ΔY126-HEK293. Мы обнаружили, что
точной культуры, так и разницы в считывании из-
концентрация GFP-p53ΔY126 меняется стохасти-
за места ее встраивания, мы повторно расклони-
чески в каждой отдельно взятой клетке от полно-
ровали линию GFP-p53ΔY126-HEK293. Все полу-
го отсутствия наблюдаемого белка к его высокой
ченные нами новые клоны также продемонстри-
концентрации через несколько часов, и наоборот
ровали разнообразие концентрации белка GFP-
(рис. 2).
БИОФИЗИКА том 67
№ 3
2022
СТОХАСТИЧНОСТЬ ЭКСПРЕССИИ БЕЛКА p53
543
Рис. 2. Изменение концентрации белка GFP-p53ΔY126 в клетках с течением времени.
Чтобы оценить кинетику экспрессии белка
рующей p53 между клетками каждой из этих куль-
GFP-p53ΔY126 в отдельных клетках исследуемой
тур, то первичные культуры клеток глиомы чело-
нами линии GFP-p53ΔY126-HEK293, мы отобра-
века демонстрируют заметную вариабельность по
ли на сортере клетки с низким («темные») и высо-
количеству кодирующей p53 РНК, от полного от-
ким
(«яркие») уровнем экспрессии GFP-
сутствия до ее значительной концентрации в каж-
p53ΔY126. Отсортированные темные клетки
дой конкретной клетке (рис. 4).
(рис. 3б) были помещены в питательную среду
при температуре 37°С. Нами было отмечено, что
Также была обнаружена зависимость количе-
после 17 ч культивирования появляются яркие
ства мРНК p53 в первичных культурах клеток
клетки, содержащие белок GFP-p53ΔY126
глиом человека от густоты посева. На редких по-
(рис. 3в). С течением времени количество флуо-
севах можно увидеть как клетки, содержащие
ресцентно окрашенных клеток увеличивалось, и
РНК, так и клетки с полным ее отсутствием, а на
спустя 72 ч соотношение «темных» и «ярких» кле-
густых посевах мы не наблюдаем значительной
ток приблизилось к распределению в исходной
разницы в количестве РНК (рис. 5).
популяции (рис. 3а,г). При дальнейшем культи-
вировании клеточной культуры в течение 30 су-
Изучение динамики восстановления уровня
ток картина распределения «темных» и «ярких»
транскрипции мРНК p53 после ее подавления раз-
клеток существенно не менялась. Однако вариа-
личными агентами. Нами было установлено, что в
бельность экспрессии белка GFP-p53ΔY126 мог-
клетках перевиваемых линий профиль мРНК p53
ла быть обусловлена как местом встраивания
не отличался от клетки к клетке при стандартных
плазмиды в генетический материал клетки, так и
условиях культивирования (рисунок 4а-в). На-
наличием мутации в гене p53ΔY126.
блюдаемое единообразие может быть обусловле-
Вариабельность мРНК, кодирующей p53 в клет-
но относительной стабильностью клеточных про-
ках первичных и перевиваемых клеточных линий.
цессов, сформировавшейся в результате длитель-
Для проверки гипотезы о стохастичности экс-
ного культивирования линий. Возможно, именно
прессии белка p53 мы проанализировали измене-
это кардинально отличает степень экспрессион-
ние концентрации кодирующей его мРНК в
ной гетерогенности в перевиваемых линиях от та-
различных клеточных линиях. Нами была ис-
ковой в первичных опухолевых линиях. Для про-
пользована методика прижизненной оценки кон-
центрации РНК с помощью зонда Smart Flare на
верки этого предположения мы решили нару-
пяти клеточных линиях. Наблюдение за концен-
шить нормальное функционирование клеток
трацией мРНК гена p53 показало наличие неко-
перевиваемых линий, влияя на различные
торых особенностей экспрессии данного гена в
внутриклеточные механизмы. Мы использовали
зависимости от условий роста и типа используе-
как агент, подавляющий общую транскрипцию в
мых культур клеток.
клетке (актиномицин Д), так и прямое специфич-
Так, для перевиваемых линий (HeLa, HT1080,
ное выключение гена с помощью РНК-интерфе-
ФЛЭЧ) и первичных культур клеток (Gl-La и Gl-
ренции. Далее с помощью конфокальной микро-
Tr) наблюдается разница в характере экспрессии
скопии визуализировали динамику восстановле-
гена p53.
ния профиля экспрессии (рис. 6). В обоих случаях
Если на перевиваемых линиях мы не отметили
мы наблюдаем стохастичное восстановление экс-
значимых различий в концентрации РНК, коди-
прессии исследуемого гена.
БИОФИЗИКА том 67
№ 3
2022
544
КОВАЛЁВ и др.
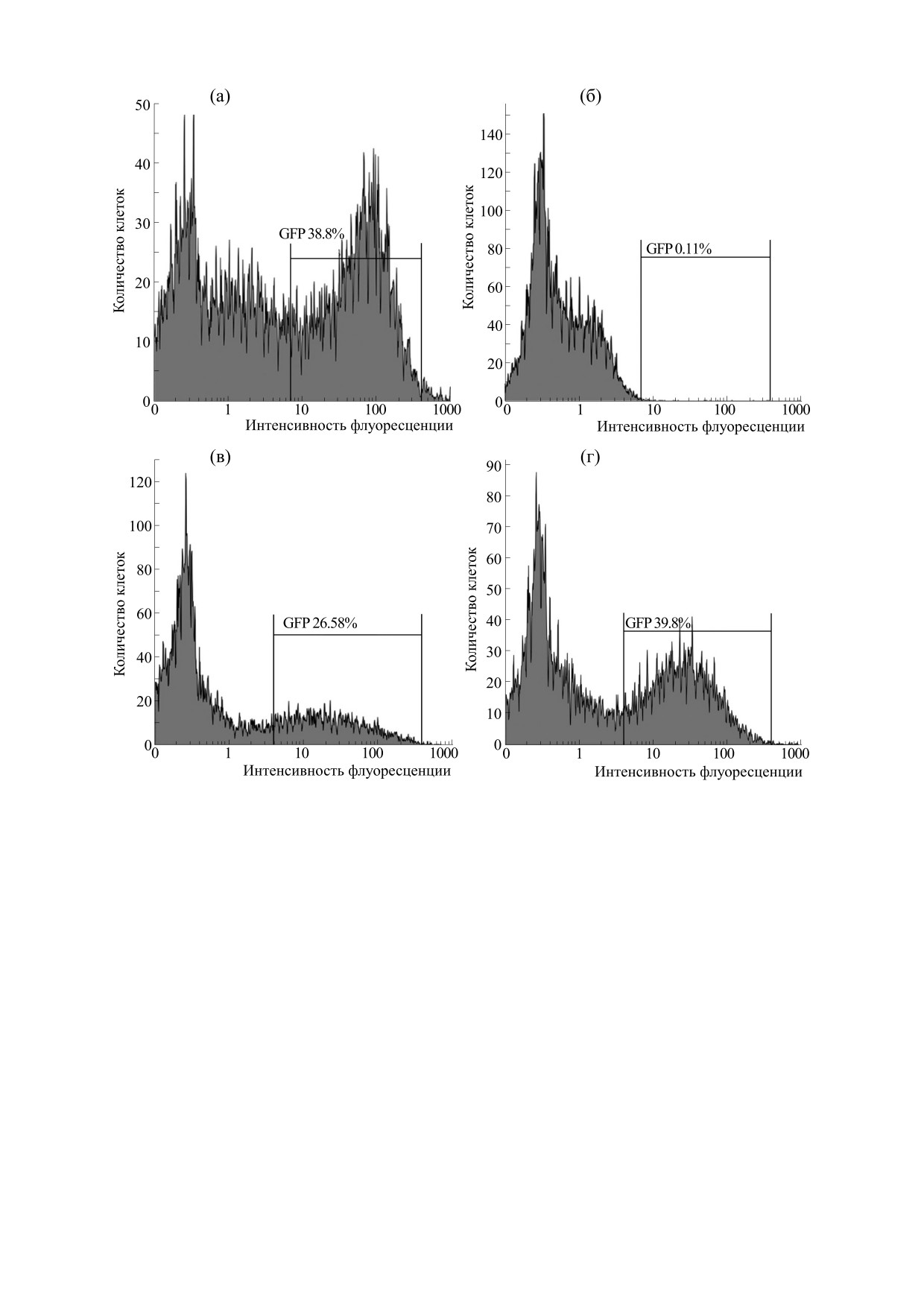
Рис. 3. Изменение уровня экспрессии GFP-p53ΔY126 в отобранной на сортере популяции клеток, изначально его не
экспрессирующих. (а) - исходная популяция клеток с различным уровнем экспрессии GFP-p53ΔY126; (б) -
отобранная на сортере популяция клеток с нулевым уровнем экспрессии GFP-p53ΔY126; (в) - уровень экспрессии
GFP-p53ΔY126 спустя 17 часов после культивирования отобранных на сортере “темных” клеток; (г) - уровень
экспрессии GFP-p53ΔY126 спустя 72 часа и более после культивирования отобранных на сортере “темных” клеток.
ОБСУЖДЕНИЕ
гулирующих экспрессию генов) [5, 12, 13]. В ре-
зультате транскрипция может носить характер че-
Клетки одного организма могут отличаться
редующихся всплесков и затуханий различной
количеством РНК, образующейся в результате
продолжительности, что приводит к неизбежной
работы одного и того же гена, а также количе-
стохастичности синтеза как мРНК, так и белка.
ством конкретных белков. Экспрессионный
Негенетическая гетерогенность, возникающая в
«шум» (случайные флуктуации экспрессии генов)
результате множественных стохастических про-
возникает по разным причинам, как внешним,
цессов, может играть значимую роль в развитии
так и внутренним (различное количество доступ-
ных клеткам питательных веществ, разная ско-
таких важных процессов, как гибель клетки, зло-
рость перемещения белков между рецепторами в
качественная трансформация, метастазирование
разных клетках, ограниченное число молекул, ре-
и др. [14, 15].
БИОФИЗИКА том 67
№ 3
2022
СТОХАСТИЧНОСТЬ ЭКСПРЕССИИ БЕЛКА p53
545
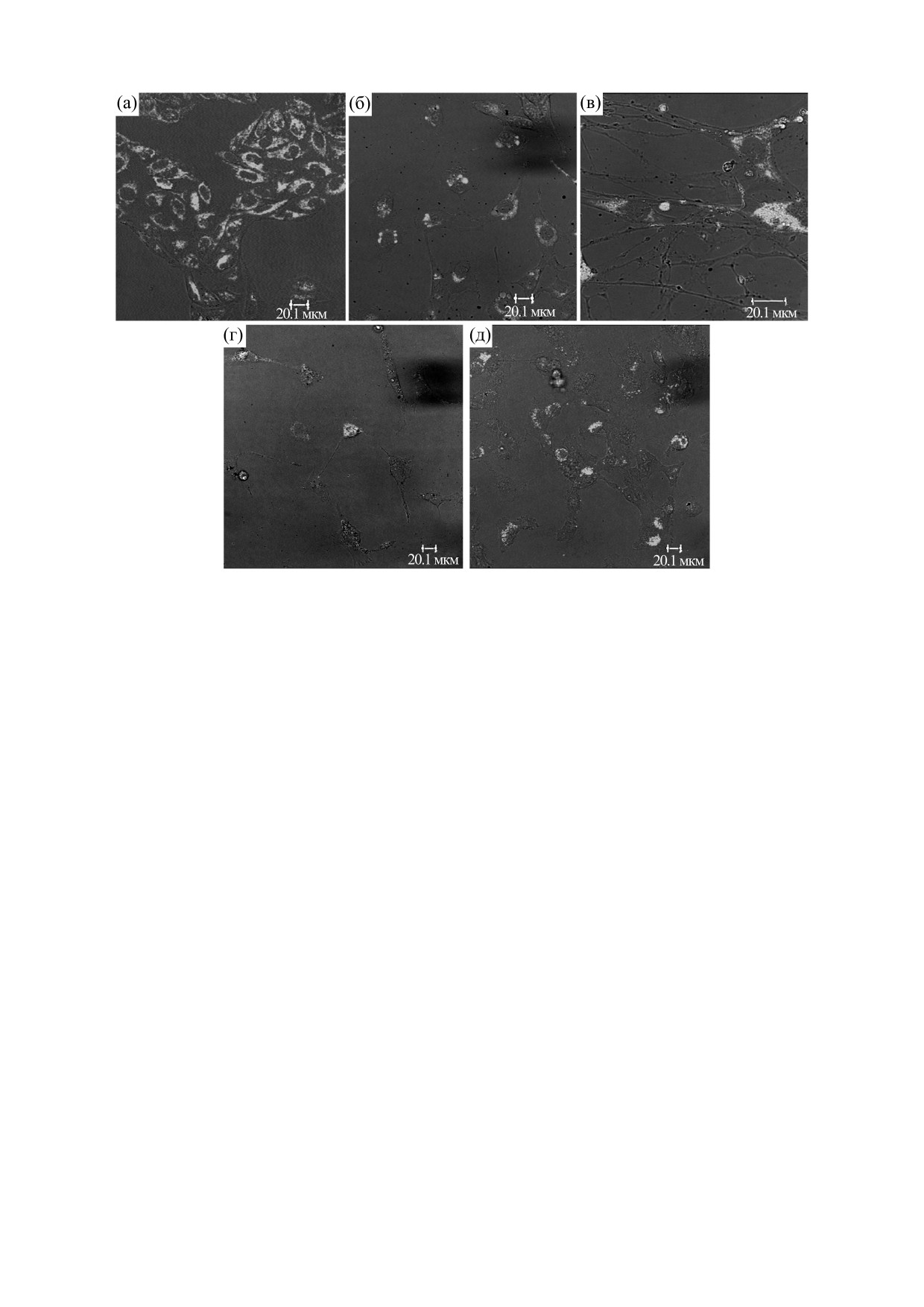
Рис. 4. Вариабельность экспрессии p53 для перевиваемых культур (HeLa - а, HT1080 - б, ФЛЭЧ - в), первичных
культур (Gl-La - г, Gl-Tr - д).
В предотвращении онкогенеза огромную роль
ствовать онкогенезу, метастазированию и сни-
играет белок p53 [16]. В ответ на активацию онко-
жать чувствительность к терапии опухолей.
генов, повреждения ДНК и другие типы стресса
Мы не отметили значимых различий в концен-
он активирует множество сигнальных путей, та-
трации РНК, кодирующей p53 между клетками
ких как остановка клеточного цикла, старение,
нормальных фибробластов человека ФЛЭЧ, как
апоптоз и др. [17]. Обычно p53 инактивируется
не было достоверных вариаций в экспрессии дан-
или через мутацию в самом гене, или посредством
ного гена и в клетках перевиваемых опухолевых
приобретения клеткой других генетических или
линий, таких как HeLa и HT1080 (рис. 4а-в). Од-
эпигенетических изменений, которые блокируют
нако мы обнаружили значительную вариабель-
функционирование p53 [18-21]. У гомозиготных
ность экспрессии p53, вплоть до полного ее отсут-
мышей с нокаутом p53 уже через 10 недель с высо-
ствия, в клетках первичных глиобластом Gl-La и
кой частотой развиваются опухоли, включая лим-
Gl-Tr (рис. 4г,д). Также мы показали, что при вве-
фомы и саркомы [22]. Считается, что для выпол-
дении в клетки HEK293 плазмиды содержащей
нения функции опухолевого супрессора необхо-
последовательность GFP-p53ΔY126, мутантный
дима постоянная экспрессия данного белка,
белок p53ΔY126 экспрессируется стохастически.
которая жестко регулируется, и в нормальных
Концентрация белка GFP-p53ΔY126 меняется с
клетках при физиологических условиях поддер-
течением времени от полного отсутствия наблю-
живается на низком уровне его отрицательными
даемого белка в клетке к высокой его концентра-
регуляторами, главным образом MDM2 и MDM4
ции через несколько часов и наоборот (рис. 3).
[23, 24]. Мутации в гене p53 или его отсутствие яв-
Можно предположить, что экспрессия генов, как
ляются признаком наследственной предрасполо-
в нормальных клетках, так и клетках перевивае-
женности к раку и часто считаются плохим про-
мых опухолевых линий, сбалансирована в боль-
гнозом для пациентов
[25,
26]. Выключение
шей степени по сравнению с клетками первич-
(инактивация) гена p53 в клетках в результате сто-
ных опухолей и клетками с введенной генетиче-
хастичности экспрессии также может способ-
ской конструкцией.
БИОФИЗИКА том 67
№ 3
2022
546
КОВАЛЁВ и др.
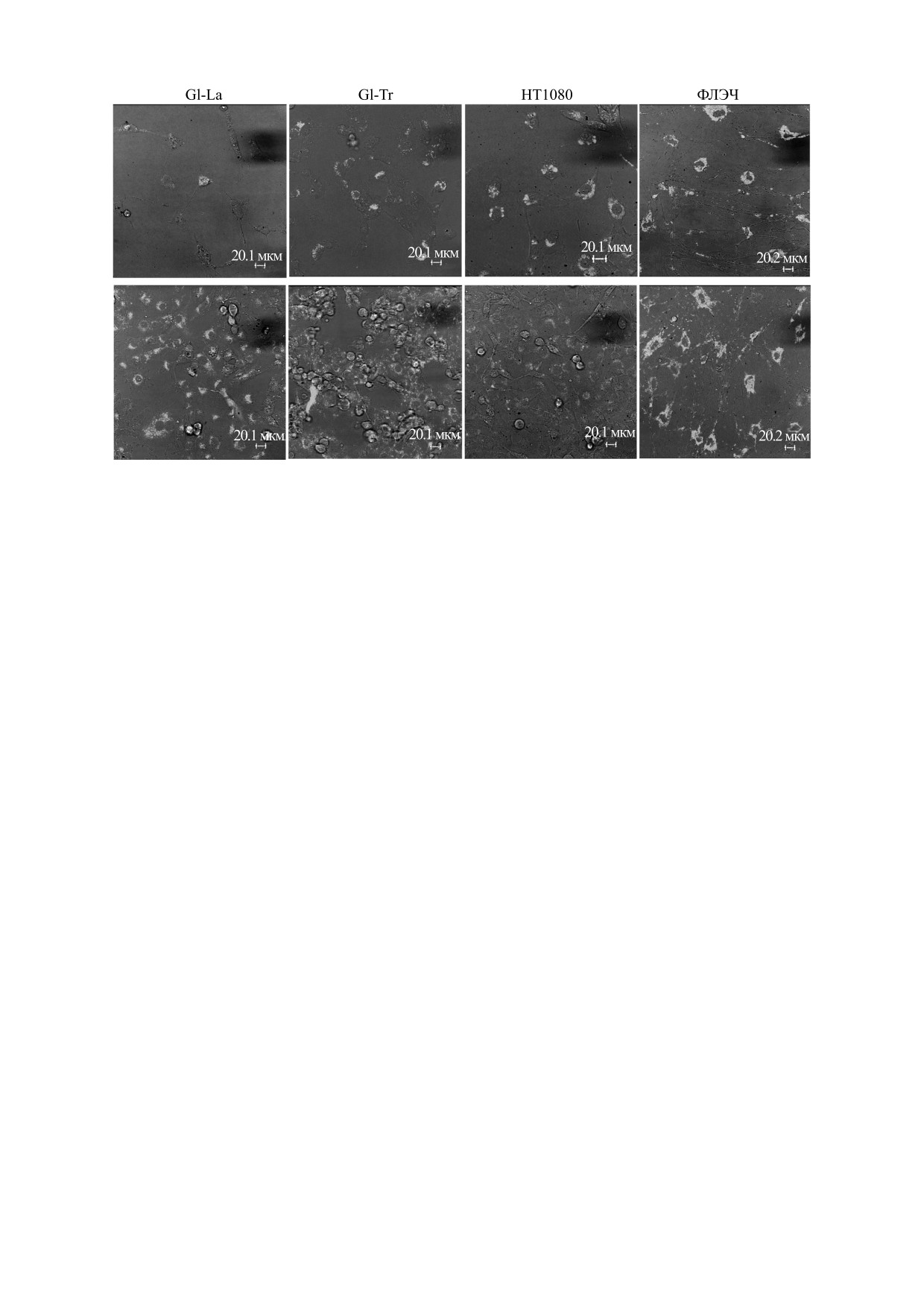
Рис. 5. Вариабельность экспрессии РНК, кодирующей p53 в клетках первичных линий (глиом Gl-La и Gl-Tr), и
перевиваемых HT1080 и ФЛЭЧ при разной густоте посева: верхний ряд - редкий посев, нижний ряд - густой посев.
Мы обнаружили, что вариабельность экспрес-
время для ее наработки и стабилизации клеточ-
сии p53 в клетках и Gl-La и Gl-Tr зависит от гу-
ных процессов, причем в разных клетках одной
стоты посева клеток (рис. 5). Так, при редком по-
популяции накопление РНК проходит стохасти-
севе наблюдаются значительные отличия в уров-
чески. Предполагается, что в регуляции стохасти-
нях экспрессии между клетками, но при
ческой экспрессии генов участвует микроРНК
дальнейшем уплотнении популяции эти разли-
[29, 30], и мы видим, что после обработки корот-
чия сглаживаются, и в монослое клетки выглядят
кой интерферирующей РНК с последующей от-
относительно однородно по уровню РНК p53.
мывкой, клеткам также требуется значительное
Эти результаты согласуются с немногочисленны-
время, чтобы ситуация стабилизировалась и пре-
ми литературными данными на эту тему. Меж-
кратились экспрессионные колебания.
клеточные взаимодействия стабилизируют и под-
Таким образом, можно заключить, что харак-
держивают относительную однородность экс-
тер экспрессии гена p53 в клетках перевиваемых
прессионных паттернов близлежащих клеток во
клеточных линий существенно не различается
время развития и дифференцировки [15, 27]. В
между клетками одной популяции. Однако при
тканях гипофиза взрослого человека клеточные
внесении возмущения и нарушении нормального
контакты обеспечивают локальную координа-
функционирования клеток наблюдается экспрес-
цию экспрессии гена пролактина [28]. Наруше-
сионная вариабельность.
ние межклеточных взаимодействий и потеря кон-
тактного торможения ответственны за повышен-
Раковые клетки характеризуются дестабили-
ный шум экспрессии, который может быть
зированным паттерном экспрессии генов [6, 31].
Кроме того, экспрессия некоторых генов ради-
причиной возникновения и прогрессии опухоли
[6, 15].
кально отличается на разных стадиях прогрессии
опухоли, что также может вносить важный вклад
Профиль экспрессии гена p53 не отличался
в увеличение негенетической гетерогенности по-
между клетками перевиваемых линий. Однако
пуляции опухолевых клеток [32]. Степень гетеро-
при нарушении нормального функционирования
генности первичных глиобластом, у которых на-
клеток, а именно подавлении транскрипции ак-
блюдаются субпопуляции с различными тран-
тиномицином Д или с использованием РНК-ин-
скрипционными профилями [33], существенно
терференции с дальнейшей заменой среды на
влияет на результаты лечения. Мы показали, что
полную, мы отметили стохастическое восстанов-
первичные клетки глиобластом могут демонстри-
ление уровня экспрессии гена p53 в клетках
ровать значительное различие уровня экспрессии
(рис. 6). Вероятно, после остановки транскрип-
гена p53 (рис. 4г,д). Представленные нами ре-
ции и снижения уровня РНК клеткам требуется
зультаты позволяют предположить, что стоха-
БИОФИЗИКА том 67
№ 3
2022
СТОХАСТИЧНОСТЬ ЭКСПРЕССИИ БЕЛКА p53
547
Рис. 6. Восстановление транскрипции РНК после прекращения воздействия актиномицином D (а) и siRNA (б).
стичность экспрессии p53 в первичных опухоле-
СПИСОК ЛИТЕРАТУРЫ
вых клетках может обуславливать их общую
1. K. Mitosch, G. Rieckh, and T. Bollenbach, Cell Syst.
нестабильность и влиять на опухолевую транс-
4 (4), 393 (2017).
формацию и метастазирование, а также, возмож-
2. A. Raj and A. van Oudenaarden, Cell 135 (2), 216
но, чувствительность к гамма- и химиотерапии,
(2008).
поскольку известно, что экспрессионный шум в
3. K. H. Kim and H. M. Sauro, PLoS Comput. Biol. 8
опухолевых клетках усиливается при низких до-
(1), e1002344 (2012).
зах ионизирующей радиации [34] и способствует
4. М. В. Головкин, Ю. Д. Нечипуренко и Г. В. Гур-
лекарственной устойчивости [35].
ский, Биофизика 54, 581 (2009).
5. P. Dong and Z. Liu, Open Biol. 7 (5), 170030 (2017).
6. J. P. Capp, Int. J. Cancer 140 (11), 2408 (2017).
ФИНАНСИРОВАНИЕ РАБОТЫ
7. A. J. Levine and M. Oren, Nat. Rev. Cancer 9, 749
Работа выполнена в рамках государственного
(2009).
задания по теме «Изучение молекулярных и кле-
8. A. F. Ramos, J. E. Hornos, and J. Reinitz, Phys. Rev.
E. Stat. Nonlin. Soft Matter Phys. 91 (2), 020701
точных компонентов патогенеза социально-зна-
(2015).
чимых заболеваний для разработки методов ран-
9. N. T. Ingolia and A. W. Murray, Curr. Biol. 17 (8), 668
ней диагностики и лечения» (регистрационный
(2007).
№ 121060200125-2).
10. V. S. Burdakov, R. A. Kovalev, R. A. Pantina, et al.,
Cell Tissue Biol. 12 (1), 20 (2018).
КОНФЛИКТ ИНТЕРЕСОВ
11. E. M. Makarov, T. A. Shtam, R. A. Kovalev, et al.,
PLoS One 12, e0185126 (2017).
Авторы заявляют об отсутствии конфликта
12. H. Xu, L. A. Sepulveda, L. Figard, et al., Nat. Meth-
интересов.
ods 12, 739 (2015).
13. C. J. Zopf, K. Quinn, J. Zeidman, and N. Maheshri,
PLoS Comput. Biol. 9, e1003161 (2013).
СОБЛЮДЕНИЕ ЭТИЧЕСКИХ СТАНДАРТОВ
14. P. W. S. Hill, R. Amouroux, and P. Hajkova, Genom-
Настоящая работа не содержит описания ис-
ics 104, 324 (2014).
следований с использованием людей и животных
15. L. Ho and G. R. Crabtree, Nature 463, 474 (2010).
в качестве объектов.
16. K. H. Vousden and C. Prives, Cell 137, 413 (2009).
БИОФИЗИКА том 67
№ 3
2022
548
КОВАЛЁВ и др.
17. Y. Liang, J. Liu, and Z. Feng, Cell Biosci. 3, 9 (2013).
27. C. V. Harper, K. Featherstone, S. Semprini, et al., J.
18. P. Hainaut, T. Hernandez, A. Robinson, et al., Nucl.
Cell Sci. 123, 424 (2010).
Acids Res. 26, 205 (1998).
28. K. Featherstone, K. Hey, H. Momiji, et al., Elife 5,
19. R. Saldana-Meyer and F. Recillas-Targa, Epigenetics
e08494 (2016).
6, 1068 (2011).
29. M. Del Giudice, S. Bo, S. Grigolon, and C. Bosia,
20. C. Kandoth, M. D. McLellan, F. Vandin, et al., Na-
PLoS Comput. Biol. 14 (4), e1006063 (2018).
ture 502 (7471), 333 (2013).
30. L. Laurenti, A. Csikasz-Nagy, M. Kwiatkowska, and
21. C. J. Brown, S. Lain, C. S. Verma, et al., Nat. Rev.
L. Cardelli, Biophys. J. 114 (12), 3000 (2018).
Cancer 9, 862 (2009).
31. A. Brock, H. Chang, and S. Huang, Nat. Rev. Genet.
22. L. A. Donehower, M. Harvey, B. L. Slagle, et al., Na-
10, 336 (2009).
ture 356, 215 (1992).
32. G. Martello and A. Smith, Ann. Rev. Cell Dev. Biol.
23. P. Hainaut, Curr. Opin. Oncol. 7, 76 (1995).
30 (1), 647 (2014).
24. A. Leonardo, S. P. Linke, K. Clarkin, and
33. A. P. Patel, I. Tirosh, J. J. Trombetta, et al., Science
G. M. Wahl, Genes Dev. 8, 2540 (1994).
344, 1396 (2014).
25. M. Olivier, M. Hollstein, and P. Hainaut, Cold Spring
34. L. Bodgi, A. Canet, A. Granzotto, et al., Cancer Ra-
Harb. Perspect. Biol. 2 (1), a001008 (2010).
diother. 20 (4), 314 (2016).
26. D. Malkin, F. P. Li, L. C. Strong, et al. Science 250,
35. K. S. Farquhar, D. A. Charlebois, M. Szenk, et al.,
1233 (1990).
Nature Commun. 10 (1), 2766 (2019).
Stochasticity in p53 Protein Expression in Human Primary Cells
and Overpassaged Cell Lines
R.А. Kovalev, N.D. Fedorova, R.А. Pantina, Е.V. Semenova, М.V. Filatov, and E.Yu. Varfolomeeva
Petersburg Nuclear Physics Institute named by B.P. Konstantinov of National Research Centre «Kurchatov Institute», Orlova
Roscha 1, Gatchina, Leningrad Region, 188300 Russia
Stochastic gene expression can have an important impact on fate of any single cell as well as on the whole cell
population. In this work, we demonstrate significant variability in p53 expression, up to its complete absence,
in primary glioblastoma cells from human tumor specimens, that does not happen in cells of normal human
fibroblasts and tumour cell lines, such as HeLa and HT1080. At the same time, variability in p53 expression
in cells depends on the density of the cell layer. We show that when the transcription is suppressed with acti-
nomycin D or p53 gene expression is knocked down by RNA interference, the level of p53 gene expression in
cells is restored and this process proceeds stochastically.
Keywords: stochastic cell heterogeneity, expression noise, p53, primary glioblastoma cells
БИОФИЗИКА том 67
№ 3
2022