БИОФИЗИКА, 2022, том 67, № 3, с. 524-530
БИОФИЗИКА КЛЕТКИ
УДК 577.3
ТЕРМОЛЮМИНЕСЦЕНЦИЯ И ДИАГНОСТИКА СОСТОЯНИЯ
ФОТОСИНТЕТИЧЕСКОГО АППАРАТА ЛИСТЬЕВ РАСТЕНИЙ
© 2022 г. В. А. Караваев, О. А. Калмацкая, Б. В. Трубицин, А. Н. Тихонов
Физический факультет Московского государственного университета
имени М.В. Ломоносова, 119991, Москва, Ленинские горы, 1/2
E-mail: karavaev@phys.msu.ru, an_tikhonov@mail.ru
Поступила в редакцию 22.03.2022 г.
После доработки 22.03.2022 г.
Принята к публикации 28.03.2022 г.
Рассмотрены основные механизмы возникновения полос термолюминесценции фотосинтетиче-
ских объектов в интервале температур от -20 до 80°С. Приведены примеры использования метода
термолюминесценции для диагностики функционального состояния фотосинтетического аппарата
растений.
Ключевые слова: термолюминесценция, оксигенный фотосинтез, электронный транспорт.
DOI: 10.31857/S0006302922030103, EDN: AOAWKT
предварительно облученных объектов [3]. На
Явление термолюминесценции (ТЛ) нашло
первом этапе основное внимание было уделено
широкое применение в исследованиях физико-
измерениям ТЛ порошков азотистых оснований
химических свойств различных систем в физике,
(аденин), нуклеотидов и нуклеиновых кислот.
биофизике, геохимии и палеонтологии. Это явле-
ние широко используется для изучения энергети-
Эти исследования показали существование про-
цессов миграции энергии возбуждения между
ческой структуры кристаллофосфоров при созда-
нуклеотидами, предположительно защищающих
нии люминесцирующих веществ, для измерения
нуклеиновые кислоты от радиационных повре-
поглощенной дозы радиоактивного излучения,
ждений. В дальнейшем основное направление
диагностики функционального состояния фото-
синтетических систем, а также для анализа
исследований было сконцентрировано на изуче-
нии фотосинтетических объектов (листья и изо-
свойств различных минералов и археологических
лированные хлоропласты). В ходе этих исследо-
артефактов [1, 2].
ваний были разработаны оптимальные протоко-
На кафедре биофизики физического факуль-
лы регистрации ТЛ, позволяющие следить за
тета МГУ имени М.В. Ломоносова исследования
функциональным состоянием фотосинтетиче-
ТЛ биологических объектов начались в середине
ского аппарата растений, произрастающих в раз-
60-х годов XX века. Эти работы были иницииро-
ных условиях и подвергнутых воздействию раз-
ваны профессором Л.А. Блюменфельдом. Пред-
ных физиологически активных соединений.
посылкой для них послужила гипотеза о том, что
Существенный вклад в изучение фотосинтетиче-
индуцированные светом процессы миграции и
ских систем внесли сотрудники кафедры биофи-
преобразования энергии в фотосинтетических
зики биологического факультета МГУ имени
системах могут протекать подобно полупровод-
М.В. Ломоносова [4].
никовым механизмам преобразования энергии в
В настоящей статье мы кратко рассмотрим ос-
физических системах. Исследования биологиче-
новы явления ТЛ и приведем примеры того, как
ских систем методом ТЛ активно велись на ка-
метод ТЛ можно использовать для диагностики
федре биофизики в течение нескольких десятиле-
функционального состояния фотосинтетическо-
тий А.К. Кукушкиным, М.К. Солнцевым и их
го аппарата растений.
учениками. М.К. Солнцев сконструировал высо-
кочувствительную экспериментальную установ-
ку, позволившую регистрировать сравнительно
ОСНОВЫ ЯВЛЕНИЯ
слабое свечение, возникающее при нагревании
ТЕРМОЛЮМИНЕСЦЕНЦИИ
Сокращения: ТЛ - термолюминесценция, ФС II - фотоси-
Термолюминесценция - это свечение, возни-
стема II, МИФ - медленная индукция флуоресценции.
кающее при нагревании объектов, предваритель-
524
ТЕРМОЛЮМИНЕСЦЕНЦИЯ И ДИАГНОСТИКА
525
но охлажденных и освещенных при пониженной
тра ФС II (Р+680) является, в конечном итоге,
температуре. При освещении вещества в нем мо-
кислородвыделяющая система. Система выделе-
гут возникать носители разноименных зарядов
ния кислорода может находиться в одном из пяти
(электроны и «дырки»), которые локализуются и
состояний: S0, S1, S2, S3 и S4. Эти состояния соот-
стабилизируются на центрах захвата («ловуш-
ветствуют состояниям ионов марганца различной
ках»). Рекомбинация разноименных зарядов мо-
степени окисленности. В результате четырех-
жет сопровождаться свечением. Для высвобожде-
кратного взаимодействия с ионами марганца раз-
ния электронов и «дырок» из «ловушек» необхо-
лагаются две молекулы воды.
дима дополнительная энергия, которую можно
сообщить либо путем нагревания объекта, либо
Выделение молекулы кислорода происходит
при его освещении инфракрасным светом [5, 6].
при переходе S4 → S0, для которого свет не требу-
Если предварительно освещенный объект под-
ется.
вергнуть нагреву в темноте, то, достигнув темпе-
ратуры, при которой тепловая энергия становит-
В работах М.К. Солнцева с соавторами [12-16]
ся сравнимой с энергией активации (энергия, не-
ТЛ листьев растений регистрировали главным
образом после их охлаждения и облучения при
обходимая для высвобождения носителя заряда
-30°С. При этой температуре «заморожены» про-
из ловушки), вещество начнет светиться. С тече-
нием времени, по мере нагревания образца, все
цессы окисления вторичного хинонового акцеп-
электрон-дырочные пары рекомбинируют и све-
тора QB-, а также переходы S4 → S0, и кратковре-
чение прекращается. Изучая зависимость интен-
менное освещение образца белым светом приво-
сивности свечения от температуры, при которой
дит к накоплению электронов на акцепторной
регистрируется излучение, получают кривую ТЛ,
стороне ФС II и дырок - в ее донорной части. По-
которая несет в себе информацию о природе и об
сле дальнейшего замораживания до азотных тем-
энергетических характеристиках ловушек носи-
ператур (77 К) и при последующем нагревании
телей зарядов.
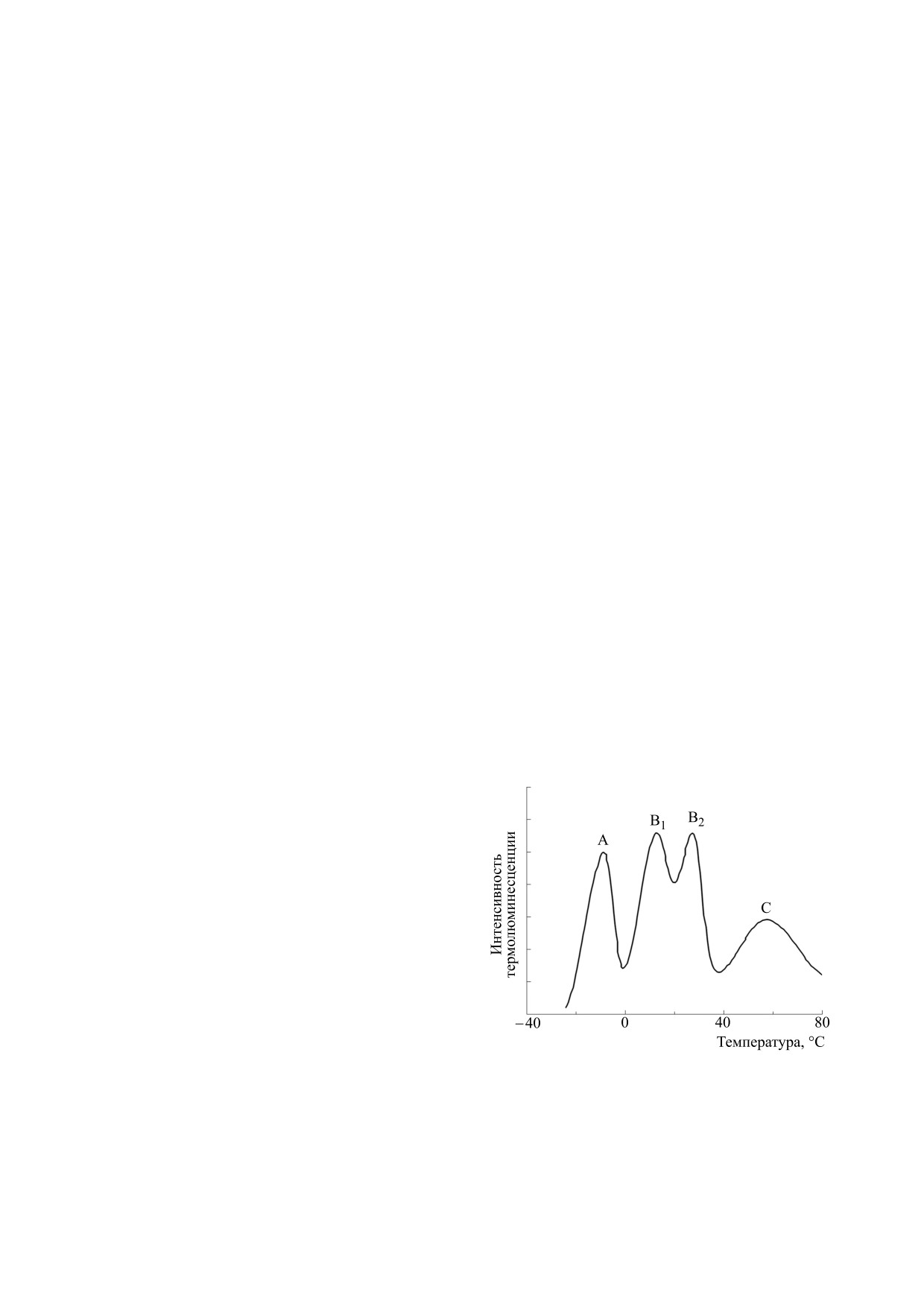
обычно наблюдались три более или менее выра-
женные полосы с максимумами в интервалах от
-20 до 0°С (полоса А), от 0 до 40°С (полоса В, со-
ИССЛЕДОВАНИЯ ФОТОСИНТЕТИЧЕСКИХ
стоящая из двух компонент В1 и В2) и от 40 до 60-
СИСТЕМ МЕТОДОМ
80°С (полоса С) [17-20] (рис. 1). В настоящее вре-
ТЕРМОЛЮМИНЕСЦЕНЦИИ
мя можно считать установленным, что полосы A
и В непосредственно связаны с функционирова-
Природа спектров термолюминесценции хлоро-
нием фотосинтетической цепи переноса электро-
пластов высших растений. Явление ТЛ фотосинте-
тических
объектов
впервые наблюдали
нов [6]. Предполагается, что полоса А возникает в
В. Арнольд и Н. Шервуд [7]. Они зарегистрирова-
основном в результате рекомбинации состояний
ли высокотемпературную термохемилюминес-
S3QA- [21], а полоса B - в результате рекомбина-
ценцию, которую связывают обычно с перекис-
−
ции S3QB- и S2Q
[6]. Существует, однако, пред-
ным окислением липидов и которая не относится
положение, что полоса A имеет двухкомпонент-
непосредственно к фотосинтезу. Компоненты,
связанные с фотосинтезом, были обнаружены
позднее (см., например, работы [8, 9]). Основным
источником свечения, испускаемого хлоропла-
стами - энергопреобразующими органеллами
растительной клетки - являются возбужденные
молекулы хлорофилла. После охлаждения образ-
ца и его освещения при низкой температуре и при
последующем нагреве происходят процессы об-
ратного переноса «дырок» от кислородвыделяю-
щей системы и электронов с хиноновых акцепто-
ров фотосистемы II (ФС II); их рекомбинация со-
провождается излучением запасенной энергии
[10, 11].
При возбуждении реакционных центров про-
исходит быстрое разделение зарядов с переносом
электрона на первичный акцептор феофитин. Да-
лее электрон переносится на первичный хиноно-
вый акцептор QA, а затем на двухэлектронный
Рис. 1. Схематическое изображение характерных по-
вторичный хиноновый акцептор QB. Донором
лос термолюминесценции фотосинтезирующих объ-
электронов для окисленного реакционного цен-
ектов.
БИОФИЗИКА том 67
№ 3
2022
526
КАРАВАЕВ и др.
ный состав; ее низкотемпературный компонент
ствовала. Очевидно, это было связано с тем, что
предварительное освещение образца проводили
вызван рекомбинацией состояний S4QA-, а высо-
при 0°С, когда еще не «заморожены» процессы
котемпературный - рекомбинацией состояний
−
окисления Q− и переходы S4 → S0.
S3Q
[20]. Выделяют еще пик Q (гербицидный,
или, в других обозначениях, G-пик), наблюдае-
В ряде работ, выполненных на физическом
мый в интервале от 2 до 10°С при блокировании
факультете МГУ имени М.В. Ломоносова, была
переноса электронов от QA к QB; предполагается,
изучена ТЛ листьев растений в условиях различ-
что эта полоса связана в основном с рекомбина-
ной фотосинтетической активности. В опытах с
−
растениями пшеницы, пораженными мучнистой
цией состояний S2Q
[6].
росой, было установлено, что изменения фото-
Полоса В состоит из двух компонент: В1 и В2.
синтетической активности (скорости выделения
Разделение этих компонент происходит, если pH
О2 на свету) положительно коррелируют с изме-
суспензии хлоропластов менее 6.0; при pH в ин-
нениями показателя SА/Sобщ, где SA - светосум-
тервале 7.0-7.5 эти две компоненты наблюдаются
ма (площадь под кривой ТЛ) пика А, Sобщ - об-
при одинаковых температурах и проявляют себя,
щая светосумма ТЛ (освещение образцов в этих
как один пик В [10]. Предполагается, что за пик
опытах проводили при -30°С) [13]. Аналогичные
В1 ответственна рекомбинация состояний S3Q−,
коррелятивные взаимосвязи были установлены в
а за пик В2 - S2QB- [2]. При дальнейшем увеличе-
экспериментах с обработкой растений различны-
ми физиологически активными веществами,
нии pH (больше 8.0) величина пика В уменьшает-
причем в условиях как повышенной, так и пони-
ся. Это связано с выпадением ионов Mn, связан-
женной фотосинтетической активности [14].
ных с кислородвыделяющей системой, что при-
водит к подавлению выделения кислорода. Этот
В другой серии экспериментов, выполненных
процесс обратим, достаточно либо уменьшить
М.К. Солнцевым с коллегами, существенные из-
pH, либо добавить NaCl [11].
менения интенсивности ТЛ в области полосы А
Высокотемпературная полоса С непосред-
наблюдали у растений, выращенных в условиях
ственно с фотосинтезом не связана. Считается,
подкормки растений удобрением «Кемира люкс».
что она обусловлена хемилюминесценцией, со-
В этих экспериментах семена пшеницы высажи-
вали в пакеты с песком объемом около 0.5 л. Пе-
провождающей реакции продуктов, которые об-
сок перед высаживанием семян обрабатывали до
разуются в результате деструкции мембран хло-
ропластов при замораживании [22]. Наличие или
полного смачивания либо водой, либо раствора-
отсутствие полосы С является определенным те-
ми удобрения «Кемира люкс» (изготовитель «Ке-
стом на устойчивость растений к тем или иным
мира Агро», Московская обл.) в пропорции 1 г
удобрения на 200 мл воды. Растения выращивали
неблагоприятным факторам среды: чем менее
в лабораторных условиях, а измерения ТЛ прово-
интенсивна полоса С, тем шире диапазон устой-
чивости растений к неблагоприятным воздей-
дили через три недели после посадки. Согласно
ствиям.
инструкции удобрение «Кемира люкс» содержит
следующие элементы: 32% азота, 20.6% фосфора,
Влияние физиологического состояния растений
27.1% калия, 0.1% железа, 0.02% бора, 0.01% меди,
на спектры термолюминесценции. Характеристики
0.1% марганца, 0.002% молибдена, 0.01% цинка.
ТЛ листьев и хлоропластов весьма чувствительны
Применение удобрения «Кемира люкс» приводи-
к изменению физиологического состояния расте-
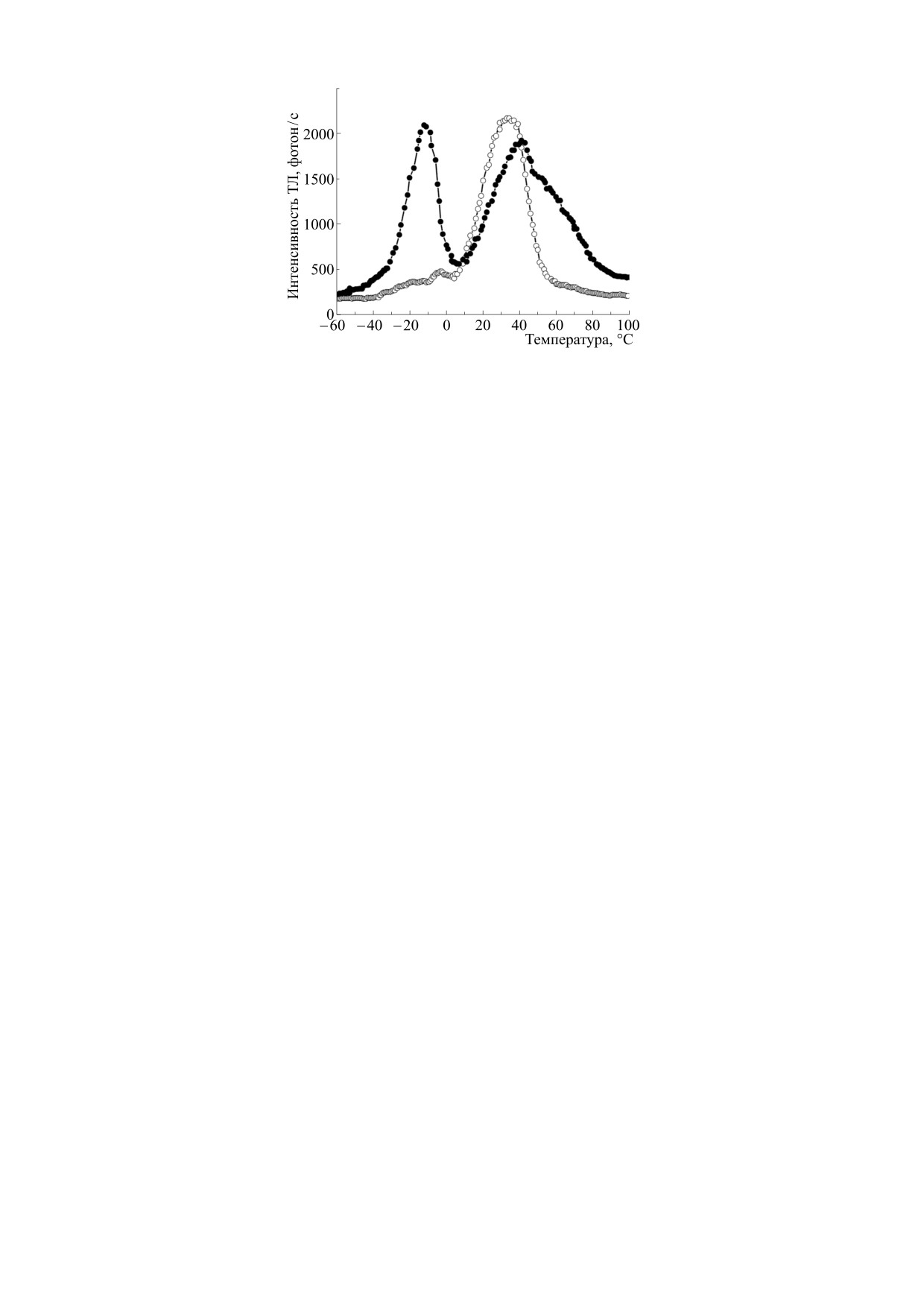
ло к значительному «разгоранию» пика А (рис. 2),
ний [2]. Рассмотрим, например, как изменяются
связанному, как предполагается, с увеличением
спектры ТЛ листьев пшеницы при различных ми-
фотосинтетической активности проростков пше-
неральных подкормках растений, которые были
ницы в условиях минеральной подкормки. Вме-
подробно изучены в работе [12]. В этих опытах
сте с тем, в этом случае наблюдалось незначи-
растения выращивали в сосудах с почвой, а поли-
тельное увеличение интенсивности высокотем-
вали их либо водопроводной водой, либо водой с
пературной ТЛ (выше 50°С), в области полосы С,
добавлением азотсодержащих, калийных или
что, очевидно, свидетельствует о некотором ухуд-
фосфорных солей. При всех типах подкормки на-
шении структурно-функциональных характери-
блюдалось увеличение показателя SB/Sобщ, где
стик мембран хлоропластов. Этот пример нагляд-
SB - светосумма (площадь под кривой) ТЛ в тем-
но иллюстрирует возможности использования
пературном диапазоне от 0 до 40°С, а Sобщ - свето-
метода ТЛ для подбора оптимальных с точки зре-
сумма ТЛ в диапазоне от 0 до 80°C. Этот эффект
ния влияния на фотосинтетический аппарат рас-
объясняется, с одной стороны, увеличением ко-
тений норм расхода препаратов.
личества QB-, накопленных к моменту регистра-
В ряде работ В.А. Караваева и М.К. Солнцева с
ции ТЛ, а с другой, - уменьшением светосуммы
соавторами [15, 16, 23, 24] было проведено ком-
полосы С. Полоса А в этих экспериментах отсут-
плексное исследование люминесцентных показа-
БИОФИЗИКА том 67
№ 3
2022
ТЕРМОЛЮМИНЕСЦЕНЦИЯ И ДИАГНОСТИКА
527
Рис. 2. Характерные кривые термолюминесценции контрольных (светлые кружки) и обработанных препаратом «Ке-
мира Люкс» (темные кружки) проростков пшеницы.
телей растений, обработанных различными фи-
обусловлена хемилюминесценцией продуктов,
зиологически активными веществами. В качестве
образующихся в результате деструкции мембран
таких показателей авторы использовали относи-
при замораживании [22]. На основании этих дан-
тельные светосуммы ТЛ (параметры SА/Sобщ и
ных был сделан вывод, что устойчивость тилако-
SС/Sобщ), а также значения относительного туше-
идных мембран к стрессовым воздействиям при
поражении трипсами снижается, а после обра-
ния флуоресценции (FM - FT)/FT при регистра-
ботки амарантином - частично восстанавливает-
ции медленной индукции флуоресценции
ся [16].
(МИФ) хлорофилла а фотосинтезирующих объ-
ектов. Так, в опытах с проростками бобов, обра-
В работе [23] исследованы МИФ и ТЛ листьев
ботанных β-аминомасляной кислотой, была
сирени Syringa vulgaris и клена серебристого Acer
установлена положительная корреляция между
saccarinum, черенки которых перед посадкой бы-
изменениями показателя (FM - FT)/FT МИФ
ли обработаны индолилмасляной кислотой, цир-
коном, корневином или препаратом рибав-экст-
листьев растений, с одной стороны, и относи-
ра. В опытах с саженцами сирени зарегистриро-
тельной светосуммы полосы А термолюминес-
вано увеличение значений (FM - FT)/FT МИФ и
ценции - с другой. Наряду с этим наблюдалось
уменьшение вклада полосы С в общую светосум-
SА/Sобщ ТЛ по сравнению с контролем в последо-
му ТЛ, что свидетельствует о положительном вли-
вательности индолилмасляная кислота → корне-
янии препарата на характеристики мембран [15].
вин → циркон, а в опытах с саженцами клена - в
В опытах с растениями огурца, пораженными
последовательности рибав-экстра → корневин →
трипсами и обработанными амарантином (азот-
→ циркон → индолилмасляная кислота. В таких же
последовательностях происходило уменьшение
содержащий алкалоид, содержащийся в листьях и
значений SС/Sобщ. Полученные результаты сви-
соцветиях амаранта), было зарегистрировано
уменьшение значений (FM - FT)/FT МИФ ли-
детельствуют о положительном влиянии обработ-
стьев огурца при поражении трипсами (на 30-
ки черенков исследованными препаратами на
физиологическое состояние проростков сирени и
35% по отношению к контролю), что свидетель-
клена.
ствовало о снижении удельной (в расчете на хло-
рофилл) фотосинтетической активности пора-
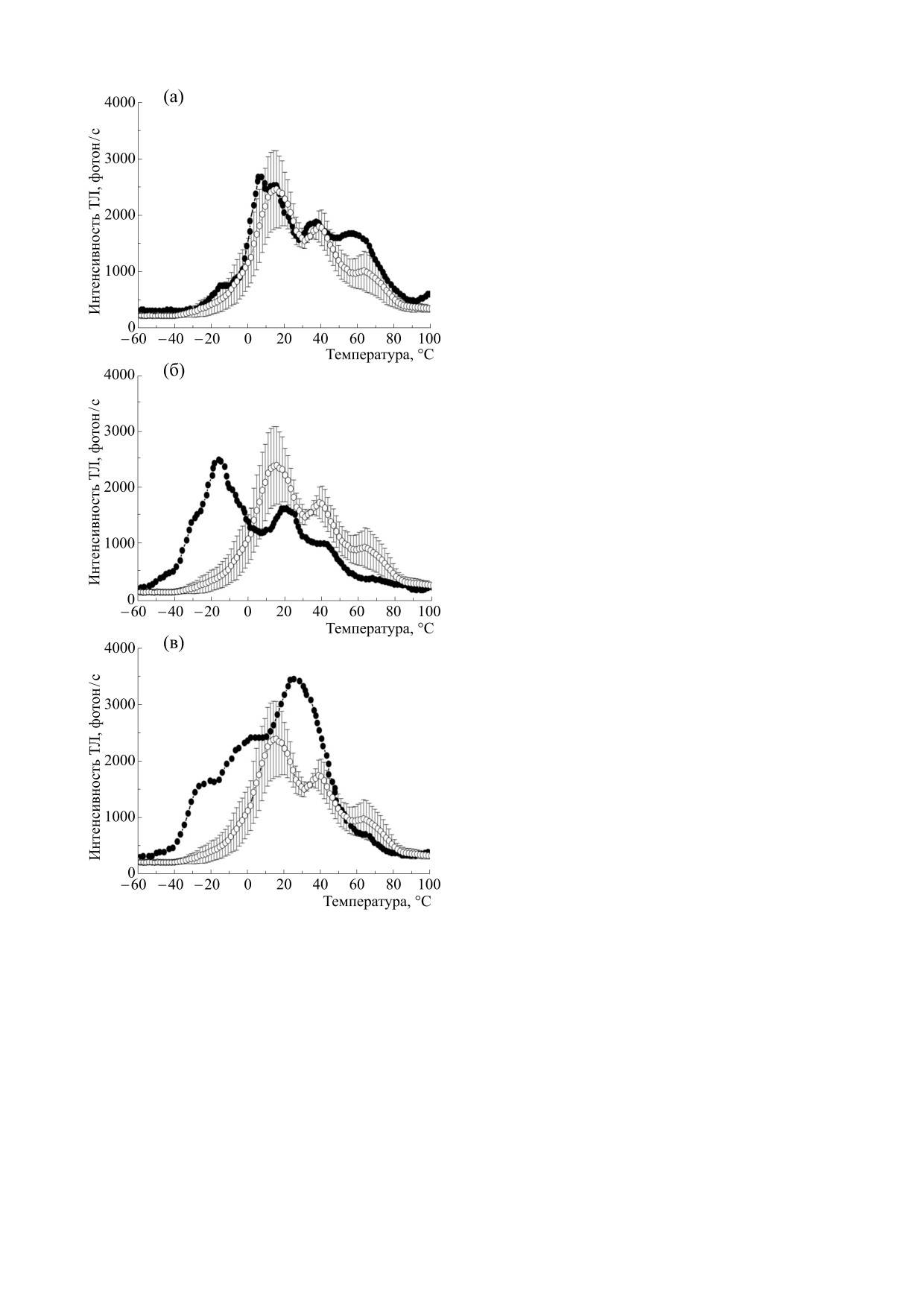
В работе [24] при обработке проростков бобов
женных листьев. После обработки листьев ама-
сверхкритическими флюидными экстрактами
рантином значения (FM - FT)/FT возрастали (до
горца сахалинского Reynoutria sachalinensis с ис-
75-80% от контроля), то есть фотосинтетическая
пользованием диоксида углерода наблюдалось
активность частично восстанавливалась. Измере-
увеличение интенсивности ТЛ в области отрица-
ния ТЛ показали, что при поражении растений
тельных температур (полоса А), но только в слу-
трипсами резко возрастает интенсивность поло-
чае, если при получении экстракта были исполь-
сы С при температурах от 40 до 80°С, а после
зованы небольшие концентрации этанола (2%)
обработки амарантином эта интенсивность
или же этанол не использовался вовсе (рис. 3).
уменьшается. Как отмечалось выше, полоса С
Расчеты показали, что в обоих этих случаях зна-
БИОФИЗИКА том 67
№ 3
2022
528
КАРАВАЕВ и др.
площадь под всей кривой ТЛ), что свидетельству-
ет об увеличении фотосинтетической активности
растений. Наиболее сильно эффект увеличения
SA/Sобщ оказался выражен при использовании
2% этанола при получении сверхкритических
флюидных экстрактов, что согласуется с данны-
ми, полученными методом МИФ. Кроме того, в
вариантах с чистым СО2 и СО2 с добавкой 2% эта-
нола наблюдалось уменьшение высокотемпера-
турной ТЛ в области полосы С, что свидетель-
ствует о повышении устойчивости мембран хло-
ропластов к неблагоприятным воздействиям [22].
При высокой (10%) концентрации этанола, ис-
пользовавшегося в качестве сорастворителя при
получении экстракта, значительно увеличивалась
интенсивность ТЛ в области полосы С, что ука-
зывает на негативное воздействие больших коли-
честв этанола на структурно-функциональные
характеристики мембран хлоропластов. Стиму-
лирующее действие экстрактов R. sachalinensis на
фотосинтетический аппарат листьев бобов может
быть связано с поступлением в клетки листа фи-
зиологически активных соединений хиноновой
природы, увеличивающих пул акцепторов элек-
трона ФС II.
В ряде работ, выполненных М.К. Солнцевым с
коллегами, методом ТЛ были изучены механизмы
действия на фотосинтетический аппарат расте-
ний ряда препаратов, обладающих гербицидным
и фунгитоксическим действием [14, 25, 26]. Боль-
шое число «фотосинтетических» гербицидов яв-
ляются ингибиторами электронного транспорта
между ФС II и ФС I. Изменения в состоянии ак-
цепторов QА и QB при обработке этими гербици-
дами приводят к соответствующим изменениям
кривых ТЛ. Эти исследования наглядно проде-
монстрировали возможности метода ТЛ для
оценки ингибирующего действия гербицидов на
первичные процессы фотосинтеза. Как отмечено
в работе [19], метод ТЛ выгодно отличается от до-
рогостоящего и трудоемкого метода, основанно-
го на регистрации радиоактивности соответству-
ющим образом меченых препаратов.
В работе [27] методами ТЛ, РАМ-флуоримет-
рии и электронного парамагнитного резонанса
было проведено сравнительное исследование фо-
Рис. 3. Характерные кривые термолюминесценции
тосинтетических характеристик листьев траде-
листьев бобов контрольных растений (светлые круж-
сканции, выращенной в условиях низкой (50-
ки) и растений, обработанных сверхкритическими
флюидными экстрактами Reynoutria sachalinensis
125 мкЭ/(м2с)) и высокой (875-1000 мкЭ/(м2с)
(темные кружки). При получении сверхкритических
освещенности. Выявлены существенные разли-
флюидных экстрактов использовали СО2
с 10% эта-
чия в соотношении светосумм полос ТЛ с макси-
(в).
нола (а), с 2% этанола (б) и чистый СО2
мумами около 0 и 25-30°С. Сделан вывод о раз-
личиях в пулах молекул пластохинона между фо-
тосистемами (ФС). Предполагается, что
чительно возрастала относительная светосумма
увеличенное количество молекул пластохинона
полосы А (показатель SA/Sобщ, где SA - площадь
на акцепторной стороне ФС II способствует эф-
под кривой ТЛ в интервале от -40 до 0°С, Sобщ -
фективному протеканию фотосинтеза в листьях
БИОФИЗИКА том 67
№ 3
2022
ТЕРМОЛЮМИНЕСЦЕНЦИЯ И ДИАГНОСТИКА
529
растений, выращенных при пониженной осве-
3.
F. Pliquett and M. K. Solntsev, Thermolumineszenz bi-
щенности.
ologischer objecte (Leipzig: VEB Georg Thieme, 1978).
Таким образом, обширные литературные дан-
4.
П. П. Нокс, П. С. Венедиктов, А. А. Кононенко
и др., Молекуляр. биология 18, 766 (1984).
ные, а также многолетние исследования, выпол-
ненные на кафедре биофизики физического фа-
5.
I. Vass and Govindjee, Photosynth. Res. 48,
117
культета МГУ, свидетельствуют о возможности
(1996).
использования метода ТЛ для оценки функцио-
6.
J.-M. Ducruet and I. Vass, Photosynth. Res. 101, 195
нальной активности фотосинтетического аппа-
(2009).
рата растений, находящихся в различных физио-
7.
W. Arnold and N. E. Sherwood, Proc. Natl. Acad. Sci.
логических условиях.
USA 43, 105 (1957).
8.
А. К. Кукушкин, Биофизика 14, 1124 (1968).
БЛАГОДАРНОСТИ
9.
I. Vass, Photosynth. Res. 76, 303 (2003).
Данная статья посвящена 100-летию со дня
10.
Y. Inoue, Biochim. Biophys. Acta 634, 309 (1981).
рождения нашего учителя, профессора Л.А. Блю-
11.
I. Vass, H. Koike, Y. Inoue, Biochim. Biophys. Acta
менфельда, который инициировал начало иссле-
810, 302 (1985).
дований методом ТЛ на кафедре биофизики фи-
12.
В. А. Караваев, М. К. Солнцев, Т. П. Юрина и др.,
зического факультета МГУ имени М.В. Ломоно-
Физиология растений 44, 20 (1997).
сова. Основной вклад в реализацию программы
13.
Т. П. Юрина, А. М. Умнов, В. А. Караваев и др.,
разнообразных исследований в этой области внес
Физиология растений 39, 270 (1992).
наш друг и коллега Михаил Константинович
Солнцев. Он оставил нам созданную им установ-
14.
M. K. Solntsev, H. P. F. Ekobena, V. A. Karavaev,
ку для изучения биологических объектов методом
et al., J. Luminescence 76&77, 349 (1998).
ТЛ и детально разработанные протоколы иссле-
15.
М. К. Солнцев, В. В. Францев, Д. Ю. Школьни-
дований фотосинтетических объектов. Остаются
ков и др., в кн. Матер. Всерос. конф. «Нетрадици-
актуальными научные открытия М.К. Солнцева в
онные и редкие растения, природные соединения и
области практических применений метода ТЛ в
перспективы их использования» (Белгород, 2006),
т. 1, сс. 379-383.
агрофизике и биофизике фотосинтеза. Неугаси-
ма светлая память о М.К. Солнцеве, которому мы
16.
М. К. Солнцев, В. В. Францев, В. А. Караваев
посвящаем эту статью.
и др., Collection of scientific papers, Faculty of agri-
culture in Ceske Budejovice. Series for Crop Sciences
Авторы выражают благодарность профессору
21, 209 (2004).
кафедры биофизики физического факультета
17.
W. Arnold and J. R. Azzi, Proc. Natl. Acad. Sci. USA
МГУ имени М.В. Ломоносова В.И. Лобышеву за
61, 29 (1968).
ценные рекомендации при написании статьи.
18.
T. Ichikawa, Y. Inoue, and K. Shibata, Biochim.
Biophys. Acta 408, 228 (1975).
ФИНАНСИРОВАНИЕ РАБОТЫ
19.
S. Demeter and Govindjee, Physiol. Plantarum 75,
Работа выполнена при финансовой поддержке
121 (1989).
Российского научного фонда (проект № 21-74-
20.
М. К. Солнцев, З. П. Грибова и В. А. Караваев,
20047).
Физиология растений 36, 686 (1989).
21.
H. Koike, Y. Siderer, T. Ono et al., Biochim. Biophys.
Acta 850, 80 (1986).
КОНФЛИКТ ИНТЕРЕСОВ
22.
М. К. Cолнцев, Журн. физ. химии 63, 1959 (1989).
Авторы заявляют об отсутствии конфликта
23.
Е. А. Кузнецова, В. А. Караваев, М. К. Солнцев
интересов.
и др., в кн. Материалы IV международной конфе-
ренции «Регуляция роста, развития и продуктивно-
сти растений» (Минск, 2005), с. 124.
СОБЛЮДЕНИЕ ЭТИЧЕСКИХ СТАНДАРТОВ
24.
С. А. Глазунова, В. А. Караваев, О. И. Покровский
Настоящая статья не содержит каких-либо ис-
и др., Сверхкритические флюиды. Теория и прак-
следований с участием людей или животных в ка-
тика, 1, 66 (2009).
честве объектов исследований.
25.
М. К. Солнцев, Л. Э. Гунар, В. Ташиш, Изв. РАН,
сер. биол. 4, 502 (1996).
СПИСОК ЛИТЕРАТУРЫ
26.
A. M. Kuznetsov, M. K. Solntsev, V. A. Karavaev,
1. К. К. Шварц, З. А. Грант, Т. К. Межс и др., Термо-
et al., in Modern fungicides and antifungal compounds
люминесцентная дозиметрия (Рига: Зинатне,
II, Ed. by H. Lyr, P.E. Russel, H.-W. Dehne, and
1968), 185 с.
H. D. Sisler, (Intercept, Andover, 1999), pp. 229-236.
2. Й. Иноу и К. Сибата, в кн. Фотосинтез, под ред.
27.
O. A. Kalmatskaya, B. V. Trubitsin, I. S. Suslichenko,
Говинджи (Мир, М., 1987), Т. 1, сс. 680-712.
et al., Photosynth. Res. 146, 123 (2020).
БИОФИЗИКА том 67
№ 3
2022
530
КАРАВАЕВ и др.
Thermoluminescence and Diagnostics of the Photosynthetic Apparatus in Plant Leaves
V.A. Karavaev, O.A. Kalmatskaya, B.V. Trubitsin, and A.N. Tikhonov
Faculty of Physics, Lomonosov Moscow State University, Leninskie Gory 1/2, Moscow, 119991 Russia
The basic mechanisms of the appearance of thermoluminescence bands of photosynthetic objects in the tem-
perature range from -20°C to 80°C are considered. Examples are given to illustrate how the thermolumines-
cence method can be used for the diagnosis of the functional state of the photosynthetic apparatus in plants.
Keywords: thermoluminescense, oxygenic photosynthesis, electron transport
БИОФИЗИКА том 67
№ 3
2022