

Почвоведение, 2023, № 9, стр. 1059-1076
Температурная чувствительность дыхания почв луговых ценозов в зоне умеренно-континентального климата: анализ данных 25-летнего мониторинга
И. Н. Курганова a, *, В. О. Лопес де Гереню a, Т. Н. Мякшина a, Д. В. Сапронов a, Д. А. Хорошаев a, В. А. Аблеева b
a Институт физико-химических и биологических проблем почвоведения РАН
142290 Московская область, Пущино, ул. Институтская, 2, Россия
b Станция фонового мониторинга, Приокско-Террасный заповедник
142200 Московская область, Данки, Россия
* E-mail: ikurg@mail.ru
Поступила в редакцию 13.03.2023
После доработки 10.05.2023
Принята к публикации 11.05.2023
- EDN: RDDRKP
- DOI: 10.31857/S0032180X23600476
Аннотация
Натурные наблюдения за дыханием почв (SR) в разных типах наземных экосистем представляются весьма актуальными, поскольку интенсивность SR характеризуется высокой временной и пространственной вариабельностью. Внутригодовая динамика SR обусловлена в значительной степени изменением гидротермических условий в течение года и часто описывается с помощью коэффициента температурной чувствительности (Q10), который во многих используемых моделях обычно имеет фиксированное значение. Целью настоящего исследования была оценка сезонной и межгодовой динамики температурной чувствительности SR в двух луговых экосистемах южного Подмосковья (умеренно-континентальный климат) на основе непрерывных 25-летних круглогодичных измерений эмиссии СО2 из почв. Луговые ценозы были сформированы на разных типах почв: дерново-подбуре (Entic Podzol (Arenic)) и серой почве (Haplic Luvisol (Loamic)). Скорость SR измеряли непрерывно с декабря 1997 г. по ноябрь 2022 г. с интервалом 7‒10 дней методом закрытых статических камер. Температурная чувствительность SR, оцененная по всей совокупности данных, имела более высокие значения на серых суглинистых почвах по сравнению с супесчаными дерново-подбурами (3.47 vs 2.59). Значения Q10 для SR в обоих типах почв в засушливые годы были в 1.2‒1.4 раза ниже, чем в годы с нормальным уровнем влажности. Межгодовая изменчивость (коэффициент вариации) значений Q10 в луговых экосистемах составила 21‒36% в зависимости от интервала температур, который принимался в расчет. В обоих луговых ценозах была обнаружена достоверная положительная корреляция между значениями Q10 в интервале температур ≥1°С и индексами влажности. Для получения более точных прогнозов баланса С в экосистемах следует применять дифференцированный подход, интегрируя в модели разные значения температурных коэффициентов для SR.
ВВЕДЕНИЕ
Дыхание почв (Soil Respiration, SR) является одним из самых значительных потоков в глобальном цикле углерода [43, 46, 56]. Вместе с тем региональные и глобальные оценки величины SR отличает самая высокая неопределенность среди других компонентов углеродного цикла [9]. Так, суммарное дыхание почв наземных экосистем нашей планеты, согласно опубликованным в разные годы данным, варьирует от 50 до 98 Гт С/год (1 Гт = 1015 г) [26, 46, 49]. Одна из последних оценок годового потока СО2 из почв планеты, полученная с использованием наиболее полной базы данных экспериментальных измерений годовой величины SR и карты растительности на основе спутниковых данных MODIS, составляет 94.3 ± ± 17.9 Гт C/год [56], что на порядок выше, чем эмиссия углекислого газа от сжигания ископаемого топлива [19]. Величины SR демонстрируют высокую пространственную [9, 27, 39] и временную неоднородность [20, 29, 31] как на уровне биомов, так и на региональном уровне. Поэтому экспериментальные наблюдения за дыханием почв в разных типах экосистем и сегодня представляются весьма актуальными.
Под термином “дыхание почв” обычно понимают выделение CO2 из почвы в атмосферу в результате двух основных процессов: разложения подстилки и органического вещества почвы за счет деятельности микро- и макроорганизмов (гетеротрофное дыхание, HR) и корневое (или автотрофное, RR) дыхание [22, 45, 60]. Являясь интегральным потоком, SR также включает выделение CO2 из почв в результате небиологических (физических и физико-химических) процессов [4, 7], но этот поток обычно незначителен [56]. Наиболее важными абиотическими факторами, влияющими на величину SR, являются температура и влажность почвы, тогда как качество и доступность субстрата являются основными биотическими факторами, которые регулируются типом растительности и ее продуктивностью [14, 44]. При натурных измерениях SR влияние температуры и других факторов трудно разделить, и их влияние неизбежно смешивается.
Зависимость величины SR от температуры воздуха или почвы, как правило, носит линейный или экспоненциальный характер. Для количественной характеристики связи между этими параметрами используют коэффициент температурной чувствительности Q10, представляющий собой величину, на которую умножается скорость SR при повышении температуры на 10°C [14, 25]. Средние значения Q10 для разных биомов колеблются незначительно: от 1.43 до 2.03, с самым высоким значением в тундре и самым низким значением в пустынях, обладая при этом существенным варьированием внутри биомов [60]. Коэффициент Q10 применяют во многих моделях углеродного цикла, используя его фиксированное значение, равное 1.5 или 2.0 для всех почв [8, 18, 38]. Вместе с тем когда пространственно неоднородные значения Q10 были интегрированы в модели вместо фиксированного значения Q10, то полученные оценки величин SR были на 40% выше. Поэтому даже небольшие неточности в значениях коэффициента Q10 могут привести к существенным погрешностям глобальных оценок почвенного дыхания и баланса С [60]. Таким образом, необходимы исследования, направленные на более глубокое понимание процессов и факторов, влияющих на температурную чувствительность SR.
При моделировании годового потока SR и баланса С в отдельных экосистемах также используют постоянное значение Q10, независимо от времени года. Однако некоторые исследования показывают, что значения коэффициента Q10 для SR не являются постоянными и меняются от сезона к сезону [11, 29, 31, 55], так как многие биотические и абиотические факторы, ответственные за температурную чувствительность SR, меняются в зависимости от времени года [24]. Принято считать, что температурная чувствительность SR отрицательно коррелирует с температурой и снижается с уменьшением влажности почвы [11, 22], так как дефицит воды может подавлять микробную активность и рост корней [14]. В то же время поступление подстилки, количество и доступность органических субстратов, активность микробного сообщества и корней растений также изменяются от сезона к сезону и могут влиять на временную (сезонную и межгодовую) динамику температурной чувствительности SR [22, 48]. Очевидно, что сезонные изменения абиотических и биотических факторов в отдельные периоды года могут по-разному влиять на динамику основных потоков, составляющих SR, – гетеротрофного, корневого и ризомикробного. Так, результаты 4-летнего эксперимента по нагреванию почвы в широколиственном лесу, расположенном в центральной Японии с прохладным умеренным климатом, показали, что влияние потепления на величину SR зависело от сезона [40]. Поток HR, представляющий один из главных компонентов SR, был более чувствителен к потеплению почвы в поздний вегетационный период по сравнению с другими сезонами. Авторы подчеркивают, что сезонные колебания воздействия потепления на SR следует учитывать при прогнозировании отклика компонентов углеродного цикла на будущие климатические изменения. В связи с этим, чтобы лучше понять механизмы неоднозначного влияния основных абиотических и биотических факторов на температурную чувствительность SR, весьма актуальными являются натурные измерения SR в различных экосистемах разных биоклиматических зон. Подавляющее большинство наблюдений за эмиссией СО2 из почв проводится в лесных или болотных экосистемах, в то время как травяные экосистемы обычно остаются в тени [6]. Вместе с тем их роль в формировании положительного баланса С на территории России довольно значительна [21].
Основная цель представляемого исследования состояла в оценке сезонной и межгодовой динамики температурной чувствительности SR на основе уникальных непрерывных 25-летних натурных измерений дыхания почвы в двух луговых ценозах южного Подмосковья, сформированных на почвах разного типа и контрастного гранулометрического состава – супесчаном дерново-подбуре и суглинистой серой почве. Задачи исследования включали (1) оценку влияния почвенных характеристик на величину Q10 и ее временную динамику, (2) поиск корреляционных связей между значениями Q10 и метеорологическими индексами, основанными на оценках температуры воздуха, количества осадков и их соотношений для различных временных интервалов в течение года. Мы предполагаем, что межгодовая изменчивость Q10 для SR в луговых экосистемах южного Подмосковья будет определяться условиями влагообеспеченности (засушливости) в период наблюдений, а гранулометрический состав почвы будет влиять на тесноту связи между температурной чувствительностью SR и метеорологическими показателями.
ОБЪЕКТЫ И МЕТОДЫ
Общая характеристика климата, растительности и почв. Исследования проводили в двух луговых ценозах, расположенных в регионе южного Подмосковья (рис. 1). Первый располагался на территории Приокско-Террасного биосферного заповедника (ПТБЗ, 54.902° N, 37.555° E), второй – вблизи г. Пущино на бывшей Опытно-полевой станции Института физико-химических и биологических проблем почвоведения РАН (ОПС И-ФХиБПП РАН, 54.834° N, 37.575° E). Регион исследований относится к зоне умеренно-континентального климата. Согласно данным многолетних метеонаблюдений, проводимых на Станции комплексного фонового мониторинга, расположенной на территории ПТБЗ (местечко Данки, Московская область; 54.902° N, 37.555° E), среднегодовая температура воздуха (±SE) в 1991‒2020 гг. в районе исследований составила 5.7 ± 0.1°С, а среднегодовое количество осадков (±SE) – 640 ± ± 20 мм (табл. 1S). Среднемесячные температуры июля и января (±SE) за этот же период составляют 18.8 ± 0.3 и –7.2 ± 0.6°С соответственно. Постоянный снежный покров в разные годы образуется в регионе, начиная с ноября до середины января, и держится, как правило, до середины апреля.
Рис. 1.
Расположение (а, c), общий вид луговых площадок (d), профили дерново-подбура (e, 1) и серой почвы (e, 2) и вид измерительных камер в вегетационный и снежный сезоны (b).
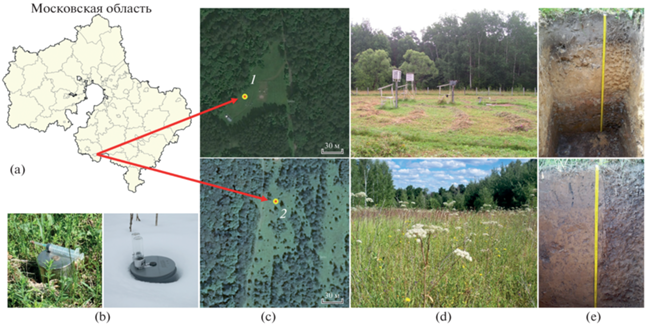
Таблица 1.
Общая характеристика почв (слой 0‒10 см) луговых ценозов (среднее ± доверительный интервал среднего, СI, p = 0.05; n = 4)
| Почва | Физическая глина*, % | НВ**, % | рНKCl | Сорг | Nобщ | С/N |
|---|---|---|---|---|---|---|
| г/кг почвы | ||||||
| Дерново-подбур | 10.3 | 35.7 ± 3.8 | 4.65 ± 0.11 | 16.6 ± 1.2 | 1.03 ± 0.04 | 16.0 |
| Серая | 36.1 | 47.3 ± 1.4 | 5.44 ± 0.35 | 20.9 ± 2.2 | 2.03 ± 0.20 | 10.3 |
Луговой ценоз на территории ПТБЗ представляет собой полидоминантную ассоциацию с преобладанием манжетки (Alchemilla sp.), фиалки трехцветной (Viola tricolor L.), трищетинника желтоватого (Trisetum flavescens (L) Beauv.), щавеля кислого (Rumex acetosa L.) и др. Почва участка ‒ дерново-подзолистая слабодифференцированная [3] на песчаных аллювиально-флювиогляциальных отложениях Приокской зандрово-аллювиальной равнины. В соответствии с Классификацией почв России 2004 г. [2] это тип дерново-подбуров или Entic Podzol (Arenic) [16]. Луговой ценоз, расположенный на территории ОПС И-ФХиБПП РАН, представляет собой старосеяный (1979 г.) разнотравно-злаковый луг с преобладанием ежи сборной (Dactylis glomerata L.), вейника наземного (Calamagrostis epigeos L.), костреца безостого (Bromopsis inermis (Leus.) Holub) и характеризуется богатым разнотравьем (Galium mollugo L., Taraxacum officinale Wigg., Hypericum perforatum L., Veronica chamaedrys L. Melampyrum nemorosum L., Chamerion angustifolium (L.) Holub.). До 2004 г. луг регулярно косили, а после прекращения сенокошения на участке стал появляться подрост из березы повислой (Betula pendula Roth). Почва участка ‒ серая лесная [3], сформирована на покровных лёссовидных породах, представленных средними и тяжелыми суглинками или легкими глинами, с преобладанием пылеватой фракции [1]. В соответствии с [2] это тип серых почв или Haplic Luvisol (Loamic) [16].
Почвы экспериментальных площадок имеют разную типовую принадлежность, контрастный гранулометрический состав (супесчаная и суглинистая) и проявляют существенные различия по другим свойствам (табл. 1). Так, при близком содержании Сорг в слое 0–10 см, содержание Nобщ в серой почве в 2 раза выше, чем в дерново-подбуре, что обусловливает существенную разницу в величине отношения С/N в верхнем 10-см слое исследуемых почв. Дерново-подбур имеет более кислую реакцию среды и существенно меньшую величину наименьшей влагоемкости по сравнению с серой почвой.
Определение дыхания почвы. Интенсивность дыхания почвы определяли непрерывно с декабря 1997 г. по ноябрь 2022 г. с интервалом 7–10 дней методом закрытых камер [37]. Измерения проводили круглогодично, в одно и тоже время суток – с 9 до 11 ч, когда текущее значение скорости SR примерно равнялось ее среднесуточному значению [30, 35]. Всего за 25-летний период наблюдений было выполнено 1126 измерений на дерново-подбуре и 1018 – на серой почве. Календарный год условно разделили на два периода: вегетационный (или бесснежный) – с мая по октябрь и холодный период (преимущественно со снежным покровом) – с ноября по апрель. Процедуры отбора проб газа в вегетационный и холодный периоды различались размерами используемых камер, их количеством (5 камер в теплый период и 3 – в холодный) и временем экспозиции [30, 37]. В бесснежный период камеры устанавливали на поверхность почвы, срезая надземную часть растений на уровне поверхности почвы. Для измерений использовали стальные (светонепроницаемые) цилиндрические камеры (диаметром 10 см и высотой 10 см). Перед началом измерений их аккуратно заглубляли в почву на 3–4 см. Концентрацию СО2 в камере определяли в течение 30 или 45 мин, отбирая газовые пробы каждые 10 мин в дерново-подбуре и каждые 15 мин – в серой почве [30, 37]. В первые 18 лет наблюдений (1997–2015 гг.) для проведения измерений в холодный снежный период на поверхность почвы заранее устанавливали стальные основания размером 32 × 32 см (с гидрозатвором), углубляя их на 20 см и используя в качестве колпаков стальные ящики размером 32 × 32 × 15 см. Чтобы исключить нарушение снежного покрова, основания при необходимости наращивались специальными секциями. Увеличение концентрации СО2 в камере измеряли в течение 135 мин с 45-минутными интервалами [37]. Начиная с ноября 2015 г., для измерения дыхания почв в холодный период года использовали стационарно установленные полихлорвиниловые основания диаметром 10 см, которые заблаговременно врезались в почву на глубину 4 см, а число таких оснований было увеличено до 5 штук на каждой мониторинговой площадке. Перед началом отбора газовых проб основания герметично закрывали крышкой, имеющей резиновую мембрану для отбора проб воздуха из камеры (рис. 1d). Пробы газа (20 см3) отбирали шприцем, доставляли в лабораторию в герметически закрытых вакуумных флаконах и в тот же день анализировали на газовом хроматографе (до 2005 г. использовали Хром-5, Чехия и затем Кристалл-2000, Россия). Скорость дыхания почв вычисляли по формуле [30]:
где SR – скорость дыхания почв, мг С/(м2 ч); С2 и С1 – конечная и начальная концентрации СО2 внутри изолятора, мг С/м3; H – высота изолятора над поверхностью почвы, м; t – время экспозиции, ч. При расчетах допускалось, что концентрация СО2 в камере нарастает линейно в первые 30 и 90 мин в теплый (май‒октябрь) и холодный (ноябрь‒апрель) периоды соответственно. Определяемый поток СО2 из почв представляет собой общее дыхание почв, т.е. сумму микробной и корневой компонент, и отчасти благодаря этому, отличается высокой пространственной вариабельностью. В отдельные сроки измерений (чаще всего в периоды оттаивания почв), величина коэффициента пространственной вариации (CVsp) могла достигать 130–150%. Среднее значение СVsp на площадках наблюдений в зависимости от года исследований составляло 12–51% в теплый период года и 21–67% – в холодный. За весь период наблюдений средняя величина CVsp в исследуемых почвах в теплый период года составляла 24–31%, а в холодный была на 10% выше – CVsp = 36–40%.Параллельно с отбором газовых проб определяли температуру почвы (Ts) на глубине 5 см. В течение 1998–2003 гг. Ts измеряли транзисторным электротермометром ТЭТ-2 (Россия). С ноября 2003 г. Ts определяли автоматическим термометром Checktemp 1 (Hanna Instruments, Германия) при отборе газовых проб, а также 6 раз в сутки термохронами iButton (США).
Оценка коэффициента температурной чувствительности Q10. Температурный коэффициент Q10 оценивали по данным еженедельных измерений SR, используя линейное регрессионное уравнение между Ln(SR) и значениями температуры почвы на глубине 5 см (Тs) [41]:
Параметры этого уравнения использовали для оценки температурного коэффициента Q10 для величины SR, который показывает во сколько раз увеличивается скорость SR при повышении температуры почвы на 10°С:
Используя полученную величину Q10, рассчитали референсное значение SR10, соответствующее скорости дыхания почвы при 10°C:
Значения Q10 и SR10 оценивались как для всего массива данных, так и дифференцированно: (1) для каждого из 25 лет наблюдений, (2) для совокупностей лет разного уровня влажности (нормальные, влажные, засушливые) и (3) для массивов данных, относящихся к отдельным календарным сезонам: зима (декабрь–февраль), весна (март–май), лето (июнь–август), осень (сентябрь–ноябрь). Для каждого набора данных были выполнены расчеты Q10 и SR10 для всего интервала Ts ($Q_{{{\text{10}}}}^{*}$ и ${\text{SR}}_{{{\text{10}}}}^{*}$), а также для Ts выше 1°C (Q10 и SR10), чтобы избежать влияния всплесков SR, в интервале температур <1°C, вызванного процессами замораживания–оттаивания. Для оценки точности линейного уравнения (2) использовали коэффициент детерминации (R2). На основании имеющихся рядов экспериментальных данных были рассчитаны “натурные” референсные величины SR10 (SR10-obs) как среднее значение экспериментально измеренных величин SR в интервале Ts = 9.0–11.0°C для всех совокупностей экспериментальных данных, описанных выше.
Анализ метеорологических данных. На основе многолетних метеоданных (среднесуточная температура воздуха (Тair, °С) и суточное количество осадков (Р, мм)), полученных на станции комплексного фонового мониторинга (Сеть наблюдений Росгидромета), расположенной на территории ПТБЗ на исследуемой луговой площадке, были оценены следующие метеорологические индексы (MI): (1) среднегодовая Тair (МАТ, °С) и годовая сумма осадков (AP, мм); (2) ST(5–8), ST(6–8), ST(6–9) и ST(5–9) – сумма среднемесячных значений Tair (°C) с мая по август, июня по август, июня по сентябрь и с мая по сентябрь соответственно; 3) SР(5–8), SР (6–8), SР(6–9) и SР(5–9) – сумма месячных сумм осадков за те же периоды, что указаны для суммы температур; 4) индексы влажности (WI): WI(5–8), WI(6–8), WI(6–9) и WI(5–9), которые представляют собой lg(SP(5–8)/ST(5–8)), lg(SP(6–8)/ST(6–8)), lg(SP(6–9)/ST(6–9)) и lg(SP(5–9)/ST(5–9)) соответственно, и 5) гидротермический коэффициент Селянинова (HTC) за летний период (июнь–август, НТС(6–8)), представляющий отношение суммы осадков (мм) за период со среднесуточной температурой воздуха выше 10°С к сумме температур за этот же период, деленное на 10 [28, 31]. Полагаем, что все эти параметры должны быть наиболее чувствительными предикторами межгодовой изменчивости значений температурного коэффициента, поскольку они охватывают различные временные интервалы в пределах вегетационного периода и будут отражать различия во влагообеспеченности в разные периоды вегетации растений. Значения индексов влажности ‒ HTC(6–8), WI(6–8) и WI(5–9) использовали также для разделения лет наблюдения на влажные, засушливые и нормальные. Если значение какого-либо из этих параметров было выше или ниже его среднего значения за период наблюдения более, чем на 1 стандартное отклонение (STD), то год относили соответственно к влажному или засушливому.
Статистическую обработку и визуализацию данных проводили в MS Exсel 2016 (пакет “Анализ данных”) и в программной среде R [42]. На графиках box plot (“ящик с усами”) показаны: среднее (крестик), медиана (черта внутри “ящика”), нижний (Q1) и верхний (Q3) квартили (“ящик”), величины Х1 = Q1 – 1.5 IQR (межквартильный размах = Q3 – Q1) и Х2 = Q3 + 1.5 IQR (“усы”), а также “выбросы” (точки на графиках), не попадающие в область между Х1 и Х2.
Перед проведением статистического анализа все независимые переменные (значения $Q_{{{\text{10}}}}^{*},$ Q10, ${\text{SR}}_{{{\text{10}}}}^{*},$ SR10 и метеоиндексы) были проверены на нормальность распределений и однородность дисперсии (критерий Шапиро–Уилка). Изменчивость величин MI, значений $Q_{{{\text{10}}}}^{*},$ Q10, ${\text{SR}}_{{{\text{10}}}}^{*},$ SR10 оценивали по следующим критериям (табл. 1S, 2S): размах варьирования (VR = max – min), коэффициент осцилляции (Cos) как отношение между VR и средним значением (Cos = VR/Mean), а также коэффициент вариации (CV, %) как отношение стандартного отклонения (STD) к среднему значению (СV = STD/Mean × 100). Для оценки влияния типа почвы и особенностей луговой растительности на средние значения $Q_{{{\text{10}}}}^{*},$ Q10, ${\text{SR}}_{{{\text{10}}}}^{*},$ SR10 использовали однофакторный дисперсионный анализ (критерий Стьюдента). Взаимосвязь между значениями Q10 и SR10 и величинами различных MI исследовали с помощью корреляции Пирсона (F-тест). Расчеты проводили с использованием уровня значимости α = 0.05, если не указано иное.
Таблица 2.
Температурная чувствительность SR луговых ценозов в разные по увлажнению годы
| Уровень увлажнения | $Q_{{{\text{10}}}}^{*},$ | ${\text{SR}}_{{{\text{10}}}}^{*},$ | R2 | n | Q10 | SR10 | R2 | n |
|---|---|---|---|---|---|---|---|---|
| Дерново-подбур | ||||||||
| Нормальный | 2.84 | 2.03 | 0.56 | 483 | 2.53 | 2.09 | 0.48 | 339 |
| Влажный | 2.94 | 2.19 | 0.63 | 220 | 2.72 | 2.26 | 0.54 | 152 |
| Сухой | 2.22 | 1.56 | 0.54 | 415 | 1.80 | 1.70 | 0.34 | 273 |
| Серая почва | ||||||||
| Нормальный | 3.69 | 1.98 | 0.59 | 440 | 2.57 | 2.27 | 0.52 | 294 |
| Влажный | 3.74 | 1.99 | 0.58 | 207 | 2.49 | 2.28 | 0.50 | 363 |
| Сухой | 3.13 | 1.59 | 0.56 | 363 | 2.20 | 1.87 | 0.50 | 266 |
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Анализ метеорологических индексов за 1998–2022 гг. В течение 25-летнего периода наблюдений метеорологические индексы сильно варьировали (рис. 2). Наибольшую изменчивость демонстрировали гидротермический коэффициент для летнего сезона HTC(6–8) и индексы, связанные с количеством осадков за вегетационный и летние периоды: SP(5–9), SP(6–8), (CV = 28–35% и Cos > 1.0) , тогда как температурные индексы ST(5–9) и ST(6–8) за этот же период характеризовались наименьшей изменчивостью с CV = 6–7% и Cos < 0.32 (табл. 1S).
Рис. 2.
Динамика метеорологических индексов за период наблюдения (А, В, С) и их статистические характеристики (а, b, c): среднее (крестик), медиана (черта), нижний (Q1) и верхний (Q3) квартили (прямоугольники), X1 = Q1 – 1.5 IQR (межквартильный размах, IQR = Q3 – Q1) и X2 = Q3 – 1.5 IQR (“усы”); а также выбросы, не попадающие в область между Х1 и Х2 (точки над “усами”). Метеорологические индексы: MAT – среднегодовая температура воздуха (°C); SТ(6–8), SТ(5–9) – сумма среднемесячных температур воздуха за летний (июнь–август) и вегетационный (май–сентябрь) периоды (°C) соответственно; SP – годовая сумма осадков (мм); SP(6–8), SP(5–9) – сумма месячных осадков за летний и вегетационный периоды соответственно; WI(6–8), WI(5–9) – индексы влажности за летний и вегетационный периоды соответственно; HTC(6–8) – гидротермический коэффициент за летний период (июнь–август). Пунктирные линии соответствуют линейным трендам, статистические характеристики которых приведены в табл. 1S.
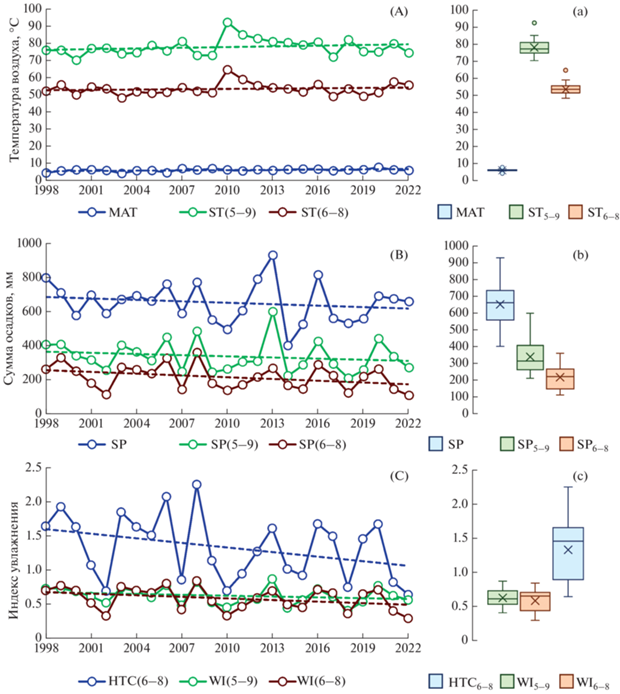
За все время проведения исследований для всех температурных индексов мы наблюдали положительные временные тренды, которые были статистически значимы (p = 0.02) только для MAT (табл. 1S). В тоже время индексы, характеризующие увлажненность, демонстрировали негативные тенденции, статистическая значимость которых не подтвердилась при p < 0.05. Следующие 5 лет были классифицированы как влажные – 1999, 2003, 2006, 2008, 2020, тогда как нижеуказанные 9 лет были классифицированы как засушливые – 2002, 2007, 2010, 2011, 2014, 2015, 2018, 2021, 2022 гг. Остальные 11 лет по уровню влажности были отнесены к нормальным годам – 1998, 2000, 2001, 2004, 2005, 2009, 2012, 2013, 2016, 2017, 2019 гг. В целом период наблюдений (1998–2022 гг.) был незначительно теплее и суше, чем период между 1991 и 2020 гг., который в настоящее время принят за климатическую норму.
Температурная чувствительность SR за весь 25-летний период наблюдений и за совокупности лет с разным уровнем увлажнения. Температурная чувствительность SR, рассчитанная для всего диапазона Ts ($Q_{{{\text{10}}}}^{*}$), в дерново-подбуре была существенно меньше, чем в серой почве: 2.59 vs 3.47 (рис. 3а, 3b). Для интервала Ts ≥ 1°C соответствующие значения Q10 были более, чем на единицу ниже, чем величины $Q_{{{\text{10}}}}^{*}$ и составили 2.19 и 2.37 для дерново-подбура и серой почвы соответственно. Референсные значения ${\text{SR}}_{{{\text{10}}}}^{*}$ и SR10 для дерново-подбура, которые были оценены по формуле (4), были близки между собой и составляли 1.86 и 1.97 г С/(м2 сут) соответственно. Для луговой экосистемы на суглинистой серой почве референсное значение SR10 для интервала температур Ts ≥ 1°C было существенно выше, чем ${\text{SR}}_{{{\text{10}}}}^{*},$ оцененное для полного интервала Ts: 2.11 vs 1.51 г С/(м2 сут).
Рис. 3.
Зависимость между Ln(SR) и температурой почвы Ts на глубине 5 см для дерново-подбура (a) и серой почвы (b) для всего ряда Ts (синяя линия) и для Ts ≥ 1°C (красная линия). SR – скорость дыхания почвы, г С/(м2 сут); Ts – температура почвы на глубине 5 см, °С; R2 – коэффициент детерминации; n – количество наблюдений; $Q_{{{\text{10}}}}^{*}$ и Q10 – температурная чувствительность дыхания почвы при полном диапазоне Ts (синий цвет) и при Ts ≥ 1°C (красный цвет), соответственно. ${\text{SR}}_{{{\text{10}}}}^{*}$ и SR10 – референсные значения SR при Тs = 10°C. a и b (± стандартная ошибка) – коэффициенты уравнения Ln(SR) = aTs + b. Все коэффициенты значимы при р $ \ll $ 0.0001.
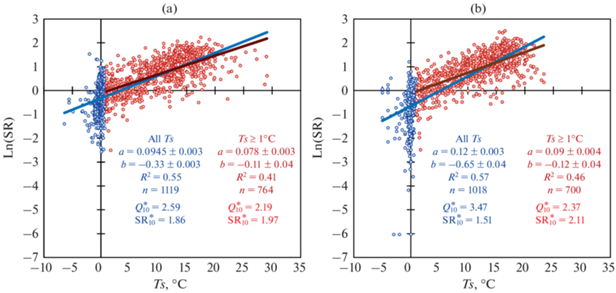
В некоторых работах также имеются указания на то, что значения Q10, основанные на полевых измерениях SR, зависят от периода наблюдений, который может охватывать весь год или ограничиваться вегетационным периодом [53]. Диапазон температур почвы для оценки значений Q10 может заметно различаться, особенно в бореальных регионах, где для почв характерны периоды промерзания. Так, в исследованных нами почвах Ts на глубине 5 см за период наблюдения могла опускаться до –5.0…–6.4°С, что свидетельствует о регулярном воздействии на почвы процессов промерзания–оттаивания. Резкий всплеск скорости SR при оттаивании почвы хорошо задокументирован [28, 32, 34], а значения Q10 в этот период могут достигать очень высоких значений (10.5–36.6) [33, 57]. Расчеты показали, что температурная чувствительность SR исследованных почв в положительном диапазоне Ts была в 1.2‒1.5 раз ниже соответствующих значений $Q_{{{\text{10}}}}^{*}$ во всем диапазоне Ts. Выполненные оценки Q10 при Ts ≥ 1°C позволили не только уменьшить их межгодовую изменчивость (рис. 3), но и в дальнейшем выявить метеорологические индексы, контролирующие величину Q10.
Существенное превышение значений $Q_{{{\text{10}}}}^{*}$ и Q10 в серой суглинистой почве, по сравнению с дерново-подбуром супесчаного гранулометрического состава, находится в полном соответствии с гипотезой СQT (carbon quality‒temperature), предложенной в работе [12]. Эта гипотеза предполагает, что величина Q10 должна увеличиваться с ростом энергии активации, необходимой для реакции разложения органического вещества почвы (SOC) [12, 13]. Следовательно, скорость деградации биохимически устойчивых органических соединений, требующих более высокой энергии активации для разложения, будет более чувствительна к увеличению температуры по сравнению с лабильным органическим веществом более низкого качества [12]. Суглинистые почвы обычно содержат больше стабильного (или связанного с минералами) органического углерода, чем песчаные почвы, которые, как правило, обогащены лабильными компонентами SOC [47]. Простым и надежным индикатором качества SOС является также отношение C/N [36, 50]. Более высокое отношение С/N в дерново-подбуре, по сравнению с серой почвой (16.0 vs 10.3, табл. 1), свидетельствует о большей подверженности SOC дерново-подбуров разложению и объясняет более низкую температурную чувствительность SR в этих почвах. В целом значения Q10 (2.19 и 2.37), полученные в настоящей работе на основании всего массива данных при Тs ≥ 1°C, близки к средним величинам Q10 = 1.81–2.60, полученным для луговых экосистем Китая [17, 36].
Выполненные в рамках настоящего исследования дифференцированные расчеты величин $Q_{{{\text{10}}}}^{*}$ и Q10 для совокупностей лет с разным уровнем увлажнения (нормальные, влажные, сухие) и для обоих интервалов температур – полного и Ts ≥ ≥ 1°C показали, что в нормальные и более влажные годы как значения $Q_{{{\text{10}}}}^{*},$ так и Q10 для SR в исследованных луговых экосистемах были близки между собой и не отличались более, чем на 0.15 единиц (табл. 2). В засушливые годы величины $Q_{{{\text{10}}}}^{*}$ и Q10 были в 1.2–1.4 раза ниже, по сравнению с годами с нормальным уровнем увлажнения, и составляли 1.56–1.87. В годы с нормальным и повышенным уровнем влажности Ts на глубине 5 см отвечала за 48–63% изменчивости SR, тогда как в засушливые годы только 34–56% дисперсии SR объяснялось колебаниями Ts на глубине 5 см. В разные по увлажнению годы температурная чувствительность SR лугового ценоза, сформированного на серой почве, была выше по сравнению с луговым ценозом на дерново-подбуре.
Расчетные значения ${\text{SR}}_{{{\text{10}}}}^{*}$ в обеих луговых экосистемах в годы с нормальным и повышенным уровнем увлажнения были близки между собой и варьировали от 1.98 до 2.19 г С/(м2 сут). Величины SR10 были выше соответствующих значений ${\text{SR}}_{{{\text{10}}}}^{*}$ и в годы с повышенным и нормальным уровнем увлажнения колебались от 2.49 до 2.72 г С/(м2 сут). В засушливые годы значения ${\text{SR}}_{{{\text{10}}}}^{*}$ и SR10 для обеих исследованных луговых экосистем были значительно ниже и изменялись от 1.56 до 1.87 г С/(м2 сут).
Таким образом, результаты показали заметное снижение значений всех расчетных коэффициентов ($Q_{{{\text{10}}}}^{*},$ Q10, ${\text{SR}}_{{{\text{10}}}}^{*}$ и SR10) в обеих почвах в засушливые годы по сравнению с нормальными по уровню увлажнения годами. Считается, что снижение влажности почвы может блокировать влияние температуры на величину SR, делая его нечувствительным к изменениям температуры [54, 55]. Температурная чувствительность SR зависит от наличия влаги, поскольку она влияет на коэффициент диффузии растворимых органических субстратов, который будет низким при недостатке влаги [60]. Низкая диффузионная способность растворимых субстратов при дефиците влаги и недостаток кислорода при высоком содержании воды снижают дыхание почвенной микробиоты, уменьшая ее температурную чувствительность [14].
Следовательно, в исследованных луговых почвах температурная чувствительность SR была обусловлена смешанным влиянием внутренних факторов, таких как содержание и состав SOC, гранулометрический состав почвы, а также уровень увлажнения почв (внешний фактор). Величины SR10 практически не зависели от внутренних особенностей изучаемых почв, но реагировали на дефицит влаги в почве. Из-за сопряженного влияния биотических и абиотических факторов значения Q10, определенные на основе измерений SR in situ, известны как “кажущиеся” (apparent) величины Q10 [13]. Поэтому использование “кажущихся” значений Q10 может привести к переоценке или недооценке влияния температуры на глобальную величину SR и исказить прогноз будущих эмиссионных потерь CO2 из почвы при глобальном потеплении.
Температурная чувствительность SR в разные календарные сезоны года. Расчеты величин $Q_{{{\text{10}}}}^{*}$ и Q10, выполненные дифференцированно для массивов данных, объединяющих отдельные календарные сезоны, показали полное отсутствие связи между Ln(SR) и Ts для летнего и зимнего периодов (Ts ≥ 1°C) и очень слабую взаимосвязь между этими параметрами в зимний период для полного ряда Тs (табл. 3). Значения $Q_{{{\text{10}}}}^{*}$ для SR в весенний сезон (март–май) в обеих исследованных луговых экосистемах были выше по сравнению с осенним периодом (сентябрь–ноябрь): 3.03–5.18 vs 2.55–3.22. Величины Q10 в весенний и осенний периоды для SR лугового ценоза на серой почве подчинялись обратной закономерности, а величины Q10 для луга на дерново-подбуре были практически равны. В весенне-осенний период колебания Ts на глубине 5 см объясняли 38–53% изменчивости SR во всем интервале температур и 29–45% при Ts ≥ 1°C.
Таблица 3.
Температурная чувствительность SR луговых ценозов в разные календарные сезоны года
| Сезон | $Q_{{{\text{10}}}}^{*}$ | ${\text{SR}}_{{{\text{10}}}}^{*}$ | R2 | n | p | Q10 | SR10 | R2 | n | p |
|---|---|---|---|---|---|---|---|---|---|---|
| Дерново-подбур | ||||||||||
| Зима | 2.84 | 1.77 | 0.04 | 252 | 0.001 | 0.65 | 0.53 | 0.008 | 43 | 0.57 |
| Весна | 3.03 | 2.25 | 0.48 | 290 | $ \ll $0.0001 | 2.39 | 2.24 | 0.32 | 167 | $ \ll $0.0001 |
| Лето | – | – | – | – | – | 1.08 | 3.12 | 0.00 | 290 | 0.43 |
| Осень | 2.55 | 1.92 | 0.38 | 288 | $ \ll $0.001 | 2.33 | 1.90 | 0.29 | 264 | $ \ll $0.0001 |
| Серая почва | ||||||||||
| Зима | 10.36 | 5.07 | 0.01 | 205 | 0.001 | 0.62 | 0.61 | 0.007 | 22 | 0.71 |
| Весна | 5.18 | 2.18 | 0.47 | 265 | $ \ll $0.0001 | 2.31 | 2.40 | 0.33 | 159 | $ \ll $0.0001 |
| Лето | – | – | – | – | – | 1.34 | 3.06 | 0.02 | 266 | 0.02 |
| Осень | 3.22 | 2.08 | 0.53 | 276 | $ \ll $0.0001 | 2.87 | 2.08 | 0.45 | 159 | $ \ll $0.0001 |
Расчетные значения ${\text{SR}}_{{{\text{10}}}}^{*}$ и SR10 в обеих луговых экосистемах для весеннего сезона были немного выше, чем в осенний период: 2.18–2.40 г С/(м2 сут) vs 1.90–2.08 г С/(м2 сут).
В литературе имеются данные, свидетельствующие о том, что температурная чувствительность SR не постоянна в течение года и меняется в зависимости от сезона [11, 40, 57]. Считается, что значения Q10 максимальны в холодный период и снижаются с повышением температуры [10, 22, 60]. Проведенные нами исследования не могли в полной мере подтвердить представления о максимальных значениях температурной чувствительности зимой и минимальной – летом, поскольку зависимости между Ln(SR) и Тs в эти периоды были чрезвычайно слабыми или отсутствовали вовсе. Тем не менее в нашем случае, значения $Q_{{{\text{10}}}}^{*}$ в весенний сезон, часто включающие значения SR из замерзших почв в ранне-весенний период, были выше, чем осенние значения $Q_{{{\text{10}}}}^{*}$, когда промерзания почв не наблюдается. Таким образом, выявленные нами различия в величинах $Q_{{{\text{10}}}}^{*}$ между весенним и осенним периодами в почвах обеих луговых экосистем находятся в соответствии с гипотезой сезонной пластичности [15], которая предполагает, что микробная активность и структура ферментов должны адаптироваться эволюционно или физиологически акклиматизироваться в ответ на сезонные изменения температуры. Часто эти процессы характеризуются более высокой температурной чувствительностью в более холодные периоды и ее снижением при повышении температуры. Однако характер сезонной динамики значений Q10 может различаться из-за большой пространственной неоднородности климатических и гидротермических условий, а также качества и доступности субстратов (SOC) разложению [38, 57]. Сильное влияние климатической зоны на сезонную динамику температурной чувствительности SR было выявлено в инкубационном эксперименте с почвами, расположенными вдоль широтного градиента на востоке Китая [58]. В сходных по составу естественных смешанных лесах температурная чувствительность SR в осенний и весенний периоды увеличивалась в направлении с севера на юг, в то время как летом значения Q10 вдоль широтного градиента были одинаковыми. Авторы предполагают, что летние значения Q10 были тесно связаны с преобладанием в микробных сообществах r-стратегов, характеризующихся высокими скоростями роста при достаточном количестве доступных (лабильных) органических субстратов. Весенние значения Q10 не зависели от состава и функций микробного сообщества, увеличиваясь с уменьшением доступности углерода с севера на юг. В то же время осенние значения Q10 определялись микробными сообществами, с преобладанием K-стратегов, что авторы объясняют наличием прайминг-эффекта, инициированного наличием свежего растительного опада [58]. Следовательно, состав микробного сообщества и продукция экзоферментов сильно зависят от сезонных изменений доступности субстрата и наличия питательных веществ, что может быть одной из причин сезонной динамики значений Q10 [40, 51, 52].
Межгодовая изменчивость температурной чувствительности и референсных значений SR в луговых экосистемах. В течение 25-летнего периода наблюдений в луговом ценозе на дерново-подбуре значения $Q_{{{\text{10}}}}^{*}$ изменялись от 1.40 до 5.42 и варьировали от 2.27 до 5.71 на лугу, сформированном на серой почве (рис. 4а).
Рис. 4.
Динамика значений $Q_{{{\text{10}}}}^{*}$ и Q10 в двух луговых экосистемах за 25-летгий период наблюдений (А, B) и их статистические характеристики (а, b): 1 – дерново-подбур, 2 – серая почва. Обозначения см. рис. 2. Пунктирные линии соответствуют линейным трендам, статистические характеристики которых приведены в табл. 2S.
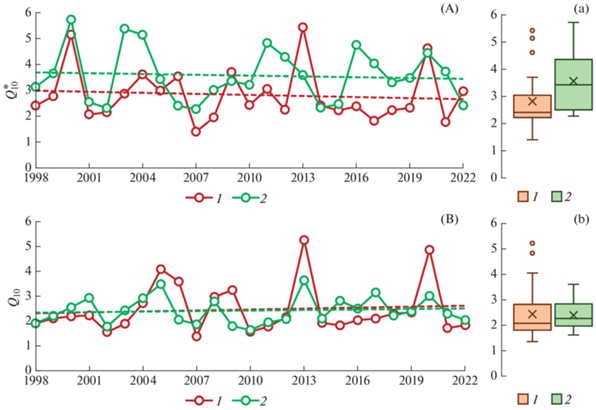
Межгодовая изменчивость значений $Q_{{{\text{10}}}}^{*}$ (CV) в этих почвах составила 36 и 29% соответственно (табл. 2S). Коэффициент осцилляции $Q_{{{\text{10}}}}^{*}$ для SR лугового ценоза на дерново-подбуре достигал 1.43, а для луга на серой лесной почве был в 1.5 раз ниже (Cos = 0.97). Средняя величина $Q_{{{\text{10}}}}^{*}$ для SR лугового ценоза на серой почве была достоверно выше, чем на дерново-подбуре (p = 0.05): 2.41 vs 3.44 (рис. 3а), а медианное значение $Q_{{{\text{10}}}}^{*}$ для дыхания серой почвы было в 1.4 раза выше, чем соответствующий показатель для дерново-подбура (рис. 4а).
Для дерново-подбура значения Q10 (интервал Ts ≥ 1°С) изменялись практически в тех же пределах, что и значения $Q_{{{\text{10}}}}^{*}$ – от 1.38 до 5.24, в то время как в серой почве диапазон межгодовой вариабельности значений Q10 был существенно уже, чем $Q_{{{\text{10}}}}^{*}$ – от 1.63 до 3.62 (рис. 4b). Коэффициенты межгодовой вариабельности и коэффициенты осцилляции для значений Q10 в дерново-подбуре были выше, а для серой почвы – чуть ниже соответствующих параметров, характеризующих варьирование $Q_{{{\text{10}}}}^{*}$ (табл. 2S). Средние значения Q10 в исследованных почвах были практически одинаковыми (2.41 и 2.46), в то время как медианное значение Q10 в дерново-подбуре было на 0.2 единицы меньше по сравнению с серой почвой: 2.12 vs 2.33.
Значения ${\text{SR}}_{{{\text{10}}}}^{*}$ в дерново-подбуре в течение 25-летнего периода наблюдений колебались от 1.30 до 3.19 г С/(м2 сут) и изменялись в более широком интервале от 0.83 до 3.74 г С/(м2 сут) – в серой почве (рис. 5a).
Рис. 5.
Динамика значений ${\text{SR}}_{{{\text{10}}}}^{*}$, SR10 и SR-obs (г С/(м2 сут)) в двух луговых ценозах за 25-летний период наблюдений (А, B, C) и их статистические характеристики (а, b, c): 1 – дерново-подбур, 2 – серая почва. Обозначения см. рис. 2. Пунктирные линии соответствуют линейным трендам, статистические характеристики которых приведены в табл. 2S.
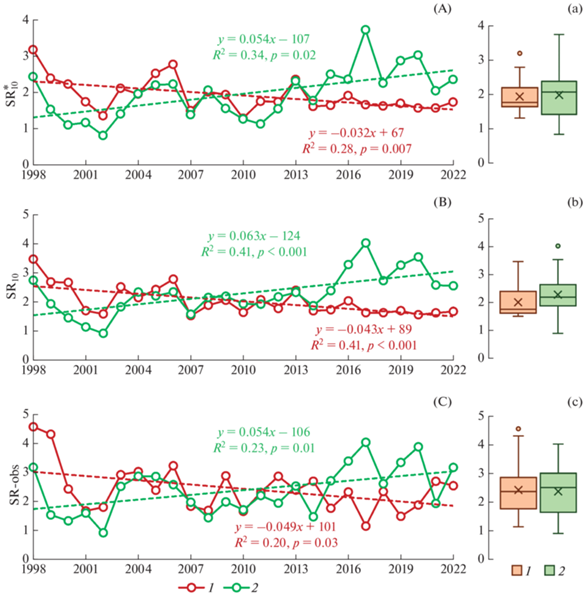
Коэффициенты межгодовой вариабельности и коэффициенты осцилляции значений ${\text{SR}}_{{{\text{10}}}}^{*}$ были существенно выше в серой почве, чем в дерново-подбуре: 23 vs 35% (СV) и 0.98 vs 1.47 (Сos) (табл. 3S). Достоверной разницы между средними значениями ${\text{SR}}_{{{\text{10}}}}^{*}$ в обеих исследованных луговых экосистемах не было выявлено, в то время как медианное значение ${\text{SR}}_{{{\text{10}}}}^{*}$ для луга на серой почве было на 0.26 г С/(м2 сут) выше, чем для луга на дерново-подбуре.
Значения SR10 (в интервале Ts ≥ 1°C) для исследованных почв изменялись в пределах, близких к соответствующим значениям ${\text{SR}}_{{{\text{10}}}}^{*}$: от 1.52 до 3.46 г С/(м2 сут) в дерново-подбуре и от 0.91 до 4.02. г С/(м2 сут) в серой лесной почве (рис. 5b). Коэффициенты межгодовой вариабельности и коэффициенты осцилляции значений SR10 также были близкими по величине к соответствующим показателям для ${\text{SR}}_{{{\text{10}}}}^{*}$ (табл. 3S), т.е. референсные значения SR10 в суглинистой серой почве демонстрировали более высокую изменчивость по сравнению с дерново-подбуром супесчаного гранулометрического состава. Достоверной разницы между средними значениями SR10 в обеих исследованных почвах не было выявлено, в то время как медианные значения SR10 в дерново-подбуре были на 0.37 г С/(м2 сут) ниже медианы SR10 в серой почве.
Значения SR10-obs изменялись в сходных пределах: от 1.14 до 4.56 г С/(м2 сут) – в дерново-подбуре и от 0.91 до 4.03 г С/(м2 сут) – в серой почве (рис. 5b). Коэффициенты межгодовой вариабельности и коэффициенты осцилляции для SR10-obs в обеих почвах были практически равными: 33–35% (CV) и 1.31–1.41 (Cos) (табл. 3S). Медианные и средние значения SR10-obs в дерново-подбуре и серой почве также были близки между собой (2.35–2.43 г С/(м2 сут)) и не отличались значимо между почвами изучаемых луговых ценозов (рис. 5c). Тесная связь (R2 = 0.49–0.79; p < 0.001) наблюдалась между значениями SR10-obs, SR10 и SR10 на протяжении всего периода наблюдений в обеих луговых экосистемах.
За 25 лет наблюдений рассчитанные и экспериментально полученные значения SR демонстрировали значимые (р < 0.05) и противоположно направленные тренды: положительные для суглинистой серой почвы и отрицательные для дерново-подбура супесчаного гранулометрического состава. Обнаруженные тенденции были наиболее выражены для SR10, объясняя более 40% вариабельности этого параметра в обеих почвах. Таким образом, наблюдаемое в регионе исследований потепление и усиление засушливости вызвало противоположные тенденции изменения SR в контрастных по текстуре почвах.
До настоящего времени межгодовая динамика температурной чувствительности SR широко не исследовалась, и основное внимание уделялось анализу пространственной изменчивости температурной чувствительности почвенного дыхания и контролирующих его факторов. Поэтому во многих моделях углеродного цикла экосистем обычно предполагается, что температурная чувствительность SR постоянна из года в год [22, 59]. Тем не менее исследования, проведенные на тибетских альпийских пастбищах в течение 4 лет (2009–2012 гг.), показали, что температурная чувствительность SR в период вегетации растений колебалась от 2.89 до 5.59 [54]. Согласно результатам 25-летних наблюдений, представленных в данном исследовании, годовая температурная чувствительность SR продемонстрировала высокую временную изменчивость с наименьшими значениями Q10 для SR в обеих исследованных почвах в засушливые годы (2002, 2007, 2010, 2022), характеризующиеся низкими индексами увлажнения. Таким образом, полученные данные подтверждают утверждение о том, что влагообеспеченность является ключевым абиотическим фактором, влияющим на температурную чувствительность дыхания почвы. В целом рост температуры ускоряет микробное дыхание за счет увеличения как активности внеклеточных ферментов, разрушающих сложные полимерные компоненты SOC, так и скорости захвата микробами растворимых субстратов [51, 52]. Ограничение влажности может подавлять микробную активность независимо от температуры, что повлечет за собой снижение температурной чувствительности SR [13]. Следовательно, имеет место смешанное влияние основных абиотических и биотических факторов на температурную чувствительность SR, оцениваемую в полевых исследованиях, о чем уже говорилось чуть выше.
Связь между значениями Q10 и SR10 и метеорологическими индексами за 25-летний период наблюдений. Для дерново-подбура супесчаной текстуры наблюдались значимые (p < 0.05) положительные корреляционные связи между величинами Q10, ${\text{SR}}_{{{\text{10}}}}^{*}$ и SR10 и всеми метеорологическими индексами, которые характеризуют режим увлажнения, а именно с суммами осадков и индексами влажности за разные периоды вегетационного сезона (рис. 6).
Рис. 6.
Корреляции между значениями $Q_{{{\text{10}}}}^{*}$, Q10, ${\text{SR}}_{{{\text{10}}}}^{*}$ и SR10 и метеорологическими индексами. Показаны только значимые корреляции (p < 0.05). $Q_{{{\text{10}}}}^{*}$ и Q10 – температурные коэффициенты дыхания почвы в полном диапазоне температуры почвы Ts и при Ts ≥ 1°С, соответственно ${\text{SR}}_{{{\text{10}}}}^{*}$ и SR10 – скорость дыхания почвы при 10°C (г С/(м2 сут), рассчитанная с использованием $Q_{{{\text{10}}}}^{*}$ и Q10. Метеорологические индексы расшифрованы в разделе Объекты и методы.
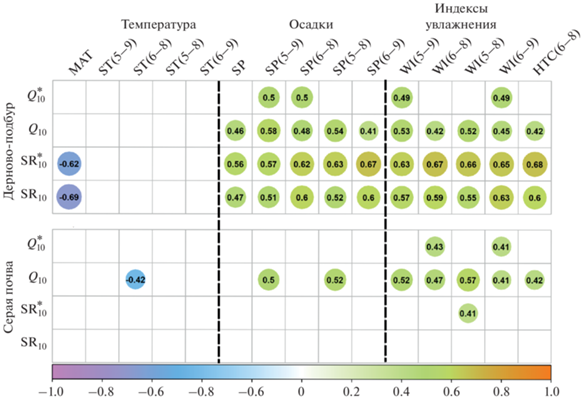
Наиболее тесные связи, объясняющие >25% дисперсии величины Q10, были выявлены с SР(5–9) и SР(5–8), WI(5–9) и WI(5–8). Величина $Q_{{{\text{10}}}}^{*}$ демонстрировала положительные связи только с SР(5–9) и SР(6–8), WI(5–9) и WI(6–8), но они были слабее и объясняли <25% дисперсии. По сравнению с Q10, величины ${\text{SR}}_{{{\text{10}}}}^{*}$ и SR10 проявляли более тесные связи с МI, и отвечали, как правило, более чем за 25% их дисперсии. Корреляция между $Q_{{{\text{10}}}}^{*}$ и Q10 и температурными индексами отсутствовала. Вместе с тем связь между ${\text{SR}}_{{{\text{10}}}}^{*}$, SR10 и МАТ была отрицательной и достаточно тесной (r = –0.62–0.69).
Для серой суглинистой почвы значимые (p < 0.05) корреляционные связи между величинами $Q_{{{\text{10}}}}^{*}$ Q10 наблюдались с более усеченным набором метеорологических индексов по сравнению с дерново-подбуром супесчаной текстуры (рис. 6). Величина Q10 положительно коррелировала со всеми индексами влажности, а также с SР(5–9) и SР(5–8) и демонстрировала отрицательную связь с ST(6–8). Величина $Q_{{{\text{10}}}}^{*}$ проявляла положительные связи только с WI(6–8) и WI(6–9). Референсные значения ${\text{SR}}_{{{\text{10}}}}^{*}$ и SR10 достоверных корреляционных связей с МI не показывали за исключением слабой положительной корреляции между ${\text{SR}}_{{{\text{10}}}}^{*}$ и WI(5–8).
Выявление факторов, контролирующих температурную чувствительность SR в луговых экосистемах, обычно основывается на метаанализе полевых или инкубационных исследований, проводимых вдоль климатического градиента [17, 36]. В целом отмечается отрицательная связь значений Q10 с температурой и положительная связь с уровнем влажности почвы. Однако до сих пор исследования по выявлению факторов, определяющих межгодовую изменчивость Q10 на локальном (экосистемном) уровне, практически отсутствуют. Такие исследования крайне необходимы для определения направления и величины обратной связи углеродного цикла отдельных экосистем с изменением климата. На основе проведенных непрерывных 25-летних измерений SR в двух луговых экосистемах показано, что основным фактором, контролирующим температурную чувствительность SR в дерново-подбуре супесчаной текстуры как в полном интервале Тs, так и при Ts ≥ 1°C, является влагообеспеченность почв в течение вегетационного периода. Между значениями $Q_{{{\text{10}}}}^{*}$ и Q10 и индексами влажности в различные по длительности отрезки вегетационного периода выявлены значимые корреляционные связи. В луговом ценозе на суглинистой серой почве значимые корреляционные связи наблюдались только между величиной Q10 при Ts ≥ 1°C и индексами влажности, но они были слабее, чем в супесчаной почве. Таким образом показано, что комплекс метеорологических показателей, контролирующих межгодовую изменчивость температурной чувствительности луговых почв в условиях умеренно континентального климата зависит от гранулометрического состава почв, который определяет их влагообеспеченность. В дерново-подбурах референсные значения скорости SR при 10°С (как ${\text{SR}}_{{{\text{10}}}}^{*},$ так и SR10) продемонстрировали сходные с Q10 зависимости, указывающие на близость контролирующих факторов и механизмов их межгодовой изменчивости. Ранее, в лесных ценозах на тех же типах почв нами были выявлены очень близкие закономерности, объясняющие временную динамику всех изучаемых параметров ($Q_{{{\text{10}}}}^{*},$ Q10, ${\text{SR}}_{{{\text{10}}}}^{*}$, SR10) условиями увлажнения и демонстрирующие тесные связями с индексами увлажненности [29].
Таким образом, проведенные исследования подтвердили гипотезу, что влагообеспеченность почв в течение вегетационного периода, определяемая метеоусловиями конкретного года исследований, была ключевым фактором межгодовой изменчивости значений Q10 в луговых экосистемах умеренного пояса, а текстура почвы влияла на тесноту связи между температурной чувствительностью SR и метеорологическими индексами. Учитывая явную тенденцию усиления засушливости климата в изучаемом регионе [5, 31] и во многих регионах мира [23], можно прогнозировать снижение температурной чувствительности дыхания почвы и предположить увеличение стока углерода в луговые экосистемы умеренно-континентального климата. В будущем моделирование скорости SR с использованием различных значений Q10 перспективно для повышения точности оценок годового потока SR, а также для уточнения региональных оценок баланса углерода. Чтобы лучше понять временную динамику температурной чувствительности SR, также необходим ежемесячный или сезонный мониторинг доступности субстрата и питательных веществ в почвах, наряду с экспериментальными оценками состава и стратегии микробных сообществ в разных почвах.
ЗАКЛЮЧЕНИЕ
Проведено обобщение 25-летних круглогодичных измерений дыхания почвы в двух луговых экосистемах в условиях умеренно-континентального климата и впервые оценена межгодовая изменчивость значений Q10 и проанализирована влияние различных факторов, определяющих сезонную и межгодовую вариабельность значений Q10. Результаты показали, что наряду с оценками Q10 для всего диапазона температур, целесообразно оценивать температурную чувствительность SR в интервале Ts ≥ 1°С чтобы избежать влияния всплесков SR, характерных для процессов промерзания-оттаивания почв. Выполненный анализ полученных данных показал, что основным фактором, контролирующим температурную чувствительность SR исследуемых почв при Ts ≥ 1°C, является влагообеспеченность в течение вегетационного периода. Об этом свидетельствуют значимые положительные корреляции между значениями Q10, количеством осадков и индексами влажности/гидротермическими показателями в различные периоды вегетационного периода. В дерново-подбуре супесчаной текстуры влияние фактора увлажненности выражено более рельефно, чем в суглинистой серой почве, обладающей большей влагоудерживающей способностью. Кроме того, обнаружили отрицательную связь величин $Q_{{{\text{10}}}}^{*}$ и Q10 со среднегодовой температурой воздуха. При существующей тенденции усиления засушливости климата можно прогнозировать снижение температурной чувствительности дыхания почв и увеличение стока углерода в луговых экосистемах умеренно-континентального климата. Для повышения точности оценок годовых потоков СО2 и моделирования баланса углерода в экосистемах целесообразно использовать дифференцированный подход с различными значениями Q10 в различные по увлажнению годы.
Список литературы
Алифанов В.М. Палеокриогенез и современное почвообразование. Пущино, 1995. 318 с.
Классификация и диагностика почв России. Смоленск: Ойкумена, 2004. 342 с.
Классификация и диагностика почв СССР. М.: Колос, 1977. 225 с.
Кобак К.И. Биотические компоненты углеродного цикла. Л.: Гидрометеоиздат, 1988. 247 с.
Курганова И.Н., Лопес де Гереню В.О., Аблеева В.А., Быховец С.С. Климат южного Подмосковья: современные тренды и оценка аномальности // Фундаментальная и прикладная климатология. 2017. № 4. С. 66–82. https://doi.org/10.21513/2410-8758-2017-4-66-82
Курганова И.Н., Лопес де Гереню В.О., Жиенгалиев А.Т., Кудеяров В.Н. Углеродный бюджет степных экосистем России // Докл. Академии наук. 2019. № 6. С. 732–735. https://doi.org/10.31857/s0869-56524856732-735
Смагин А.В. Газовая функция почв // Почвоведение. 2000. № 10. С. 1211–1223.
Bond-Lamberty B., Thomson A. Temperature-associated increases in the global soil respiration record // Nature. 2010. V. 7288. P. 579–582. https://doi.org/10.1038/nature08930
Bond-Lamberty B., Thomson A. A global database of soil respiration data // Biogeosciences. 2010. V. 6. P. 1915–1926. https://doi.org/10.5194/bg-7-1915-2010
Bradford M.A., Davies C.A., Frey S.D., Maddox T.R., Melillo J.M., Mohan J.E., Reynolds J.F., Treseder K.K., Wallenstein M.D. Thermal adaptation of soil microbial respiration to elevated temperature // Ecology Lett. 2008. V. 12. P. 1316–1327. https://doi.org/10.1111/j.1461-0248.2008.01251.x
Chen B., Liu S., Ge J., Chu J. Annual and seasonal variations of Q10 soil respiration in the sub-alpine forests of the Eastern Qinghai-Tibet Plateau, China // Soil Biol. Biochem. 2010. V. 10. P. 1735–1742. https://doi.org/10.1016/j.soilbio.2010.06.010
Craine J.M., Fierer N., McLauchlan K.K. Widespread coupling between the rate and temperature sensitivity of organic matter decay // Nature Geoscience. 2010. V. 12. P. 854–857. https://doi.org/10.1038/ngeo1009
Davidson E.A., Janssens I.A. Temperature sensitivity of soil carbon decomposition and feedbacks to climate change // Nature. 2006. V. 7081. P. 165–173. https://doi.org/10.1038/nature04514
Davidson E.A., Janssens I.A., Luo Y. On the variability of respiration in terrestrial ecosystems: moving beyond Q10 // Global Change Biology. 2006. V. 2. P. 154–164. https://doi.org/10.1111/j.1365-2486.2005.01065.x
Drake J.E., Giasson M.-A., Spiller K.J., Finzi A.C. Seasonal plasticity in the temperature sensitivity of microbial activity in three temperate forest soils // Ecosphere. 2013. V. 6. P. 77. https://doi.org/10.1890/ES13-00020.1
FAO World reference base for soil resources 2014: international soil classification system for naming soils and creating legends for soil maps / FAO, Rome: FAO, 2014.
Feng J., Wang J., Song Y., Zhu B. Patterns of soil respiration and its temperature sensitivity in grassland ecosystems across China // Biogeosciences. 2018. V. 17. P. 5329–5341. https://doi.org/10.5194/bg-15-5329-2018
Foereid B., Ward D.S., Mahowald N., Paterson E., Lehmann J. The sensitivity of carbon turnover in the Community Land Model to modified assumptions about soil processes // Earth System Dynamics. 2014. V. 1. P. 211–221. https://doi.org/10.5194/esd-5-211-2014
Friedlingstein P., Jones M.W., O’Sullivan M., Andrew R.M., Hauck J., Peters G.P., Peters W. et al. Global Carbon Budget 2019 // Earth System Science Data. 2019. V. 4. P. 1783–1838. https://doi.org/10.5194/essd-11-1783-2019
Giasson M.-A., Ellison A.M., Bowden R.D., Crill P.M., Davidson E.A., Drake J.E., Frey S.D. et al. Soil respiration in a northeastern US temperate forest: a 22-year synthesis // Ecosphere. 2013. V. 11. Р. 140. https://doi.org/10.1890/es13.00183.1
Golubyatnikov L.L., Kurganova I.N., Lopes de Gerenyu V.O. Estimation of Carbon Balance in Steppe Ecosystems of Russia // Izvestiya, Atmospheric and Oceanic Physics. 2023. V. 1. P. 72–86. https://doi.org/10.1134/S0001433823010048
Han M., Jin G. Seasonal variations of Q10 soil respiration and its components in the temperate forest ecosystems, northeastern China // Eur. J. Soil Biol. 2018. V. 85. P. 36–42. https://doi.org/10.1016/j.ejsobi.2018.01.001
IPCC Climate Change 2021: The Physical Science Basis. Contribution of Working Group I to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change. IPCC, Cambridge, United Kingdom and New York, NY, USA: Cambridge University Press, 2021. 3949 p.
Janssens I.A., Pilegaard K. Large seasonal changes in Q10 of soil respiration in a beech forest // Global Change Biology. 2003. V. 6. P. 911–918. https://doi.org/10.1046/j.1365-2486.2003.00636.x
Kirschbaum M.U.F. The temperature dependence of soil organic matter decomposition, and the effect of global warming on soil organic C storage // Soil Biol. Biochem. 1995. V. 6. P. 753–760. https://doi.org/10.1016/0038-0717(94)00242-S
Kudeyarov V.N. Soil Respiration and Biogenic Carbon Dioxide Sink in the Territory of Russia: An Analytical Review // Eurasian Soil Science. 2018. V. 6. P. 599–612. https://doi.org/10.1134/s1064229318060091
Kudeyarov V.N., Kurganova I.N. Respiration of Russian Soils: Database Analysis, Long-Term Monitoring, and General Estimates // Eurasian Soil Science. 2005. V. 9(38). P. 983–992.
Kurganova I., Lopes de Gerenyu V., Khoroshaev D., Blagodatskaya E. Effect of snowpack pattern on cold-season CO2 efflux from soils under temperate continental climate // Geoderma. 2017. V. 304. P. 28–39. https://doi.org/10.1016/j.geoderma.2016.09.009
Kurganova I., Lopes de Gerenyu V., Khoroshaev D., Myakshina T., Sapronov D., Zhmurin V. Temperature Sensitivity of Soil Respiration in Two Temperate Forest Ecosystems: The Synthesis of a 24-Year Continuous Observation // Forests. 2022. V. 9. P. 1374. https://doi.org/10.3390/f13091374
Kurganova I., Lopes de Gerenyu V., Rozanova L., Sapronov D., Myakshina T., Kudeyarov V. Annual and seasonal CO2 fluxes from Russian southern taiga soils // Tellus B: Chemical and Physical Meteorology. 2003. V. 2. P. 338–344. https://doi.org/10.3402/tellusb.v55i2.16724
Kurganova I.N., Lopes de Gerenu V.O., Khoroshaev D.A., Myakshina T.N., Sapronov D.V., Zhmurin V.A., Kudeyarov V.N. Analysis of the long-term soil respiration dynamics in the forest and meadow cenoses of the Prioksko-Terrasny biosphere reserve in the perspective of current climate trends // Eurasian Soil Science. 2020. V. 10. P. 1421–1436. https://doi.org/10.1134/s1064229320100117
Kurganova I.N., Lopes de Gerenyu V.O. Contribution of abiotic factors to CO2 emission from soils in the freeze–thaw cycles // Eurasian Soil Science. 2015. V. 9. P. 1009–1015. https://doi.org/10.1134/s1064229315090082
Kurganova I.N., Tipe P. The effect of freezing-thawing processes on soil respiration activity // Eurasian Soil Science. 2003. V. 9. P. 976–985
Kurganova I., Teepe R., Loftfield N. Influence of freeze-thaw events on carbon dioxide emission from soils at different moisture and land use // Carbon Balance and Management. 2007. V. 1(2). Р. 2. https://doi.org/10.1186/1750-0680-2-2
Larionova A.A., Rozanova L.N., Samoylov T.I. Dynamics of gas exchange in the profile of a gray forest soil // Soviet soil science. 1989. V. 3. P. 104–110.
Liu Y., He N., Zhu J., Xu L., Yu G., Niu S., Sun X., Wen X. Regional variation in the temperature sensitivity of soil organic matter decomposition in China’s forests and grasslands // Global Change Biology. 2017. V. 8. P. 3393–3402. https://doi.org/10.1111/gcb.13613
Lopes de Gerenyu V.O., Kurganova I.N., Rozanova L.N., Kudeyarov V.N. Annual emission of carbon dioxide from soils of the Southern Taiga soils of Russia // Eurasian Soil Science. 2001. V. 34. P. 931–944.
Meyer N., Welp G., Amelung W. The temperature sensitivity (Q10) of soil respiration: controlling factors and spatial prediction at regional scale based on environmental soil classes // Global Biogeochemical Cycles. 2018. V. 2. P. 306–323. https://doi.org/10.1002/2017GB005644
Mukhortova L., Schepaschenko D., Moltchanova E., Shvidenko A., Khabarov N., See L. Respiration of Russian soils: Climatic drivers and response to climate change // Sci. Total Environ. 2021. V. 785. P. 147314. https://doi.org/10.1016/j.scitotenv.2021.147314
Noh N.J., Kuribayashi M., Saitoh T.M., Muraoka H. Different responses of soil, heterotrophic and autotrophic respirations to a 4-year soil warming experiment in a cool-temperate deciduous broadleaved forest in central Japan // Agricultural and Forest Meteorology. 2017. V. 247. P. 560–570. https://doi.org/10.1016/j.agrformet.2017.09.002
Pavelka M., Acosta M., Marek M. V., Kutsch W., Janous D. Dependence of the Q10 values on the depth of the soil temperature measuring point // Plant and Soil. 2007. V. 1–2. P. 171–179. https://doi.org/10.1007/s11104-007-9213-9
R Core Team R: A language and environment for statistical computing / R Core Team, Vienna: R Foundation for Statistical Computing, 2021.
Raich J.W., Tufekcioglu A. Vegetation and soil respiration: Correlations and controls // Biogeochemistry. 2000. V. 1(48). P. 71–90. https://doi.org/10.1023/A:1006112000616
Reichstein M., Rey A., Freibauer A., Tenhunen J., Valentini R., Banza J., Casals P., Cheng Y. et al. Modeling temporal and large-scale spatial variability of soil respiration from soil water availability, temperature and vegetation productivity indices // Global Biogeochemical Cycles. 2003. V. 4. P. 1104. https://doi.org/10.1029/2003GB002035
Rustad L.E., Huntington T.G., Boone R.D. Controls on soil respiration: Implications for climate change // Biogeochemistry. 2000. V. 1. P. 1–6. https://doi.org/10.1023/a:1006255431298
Schlesinger W.H., Andrews J.A. Soil respiration and global carbon cycle // Biogeochemistry. 2000. V. 1. P. 7–20. https://doi.org/10.1023/a:1006247623877
Schmidt M.W.I., Torn M.S., Abiven S., Dittmar T., Guggenberger G., Janssens I.A., Kleber M., Kögel-Knabner I. et al. Persistence of soil organic matter as an ecosystem property // Nature. 2011. V. 7367. P. 49–56. https://doi.org/10.1038/nature10386
Shabaga J.A., Basiliko N., Caspersen J.P., Jones T.A. Seasonal controls on patterns of soil respiration and temperature sensitivity in a northern mixed deciduous forest following partial-harvesting // Forest Ecology and Management. 2015. V. 348. P. 208–219. https://doi.org/10.1016/j.foreco.2015.03.022
Sitch S., Friedlingstein P., Gruber N., Jones S.D., Murray-Tortarolo G., Ahlström A., Doney S.C. at al. Recent trends and drivers of regional sources and sinks of carbon dioxide // Biogeosciences. 2015. V. 3. P. 653–679. https://doi.org/10.5194/bg-12-653-2015
Sollins P., Homann P., Caldwell B.A. Stabilization and destabilization of soil organic matter: mechanisms and controls // Geoderma. 1996. V. 1. P. 65–105. https://doi.org/10.1016/S0016-7061(96)00036-5
Suseela V., Conant R.T., Wallenstein M.D., Dukes J.S. Effects of soil moisture on the temperature sensitivity of heterotrophic respiration vary seasonally in an old-field climate change experiment // Global Change Biology. 2012. V. 1. P. 336–348. https://doi.org/10.1111/j.1365-2486.2011.02516.x
Wallenstein M.D., Mcmahon S.K., Schimel J.P. Seasonal variation in enzyme activities and temperature sensitivities in Arctic tundra soils // Global Change Biology. 2009. V. 7. P. 1631–1639. https://doi.org/10.1111/j.1365-2486.2008.01819.x
Wang W., Peng S., Wang T., Fang J. Winter soil CO2 efflux and its contribution to annual soil respiration in different ecosystems of a forest-steppe ecotone, north China // Soil Biol. Biochem. 2010. V. 3. P. 451–458. https://doi.org/10.1016/j.soilbio.2009.11.028
Wang Y., Liu H., Chung H., Yu L., Mi Z., Geng Y., Jing X., Wang S., Zeng H., Cao G., Zhao X., He J.-S. Non-growing-season soil respiration is controlled by freezing and thawing processes in the summer monsoon-dominated Tibetan alpine grassland // Global Biogeochemical Cycles. 2014. V. 10. P. 1081–1095. https://doi.org/10.1002/2013GB004760
Xu M., Qi Y. Spatial and seasonal variations of Q10 determined by soil respiration measurements at a Sierra Nevadan Forest // Global Biogeochemical Cycles. 2001. V. 3(15). P. 687–696. https://doi.org/10.1029/2000GB001365
Xu M., Shang H. Contribution of soil respiration to the global carbon equation // J. Plant Physiology. 2016. V. 203. P. 16–28. https://doi.org/10.1016/j.jplph.2016.08.007
Yang L., Zhang Q., Ma Z., Jin H., Chang X., Marchenko S.S., Spektor V.V. Seasonal variations in temperature sensitivity of soil respiration in a larch forest in the Northern Daxing’an Mountains in Northeast China // J. Forestry Research. 2022. V. 3. P. 1061–1070. https://doi.org/10.1007/s11676-021-01346-4
Yang S., Wu H., Wang Z., Semenov M.V., Ye J., Yin L., Wang X., Kravchenko I., Semenov V., Kuzyakov Y., Jiang Y., Li H. Linkages between the temperature sensitivity of soil respiration and microbial life strategy are dependent on sampling season // Soil Biol. Biochem.2022. V. 172. P. 108758. https://doi.org/10.1016/j.soilbio.2022.108758
Zheng Z.-M., Yu G.-R., Fu Y.-L., Wang Y.-S., Sun X.-M., Wang Y.-H. Temperature sensitivity of soil respiration is affected by prevailing climatic conditions and soil organic carbon content: A trans-China based case study // Soil Biol. Biochem. 2009. V. 7(41). P. 1531–1540. https://doi.org/10.1016/j.soilbio.2009.04.013
Zhou T., Shi P., Hui D., Luo Y. Global pattern of temperature sensitivity of soil heterotrophic respiration (Q10) and its implications for carbon-climate feedback // J. Geophysical Research: Biogeosciences. 2009. V. G2 (114). https://doi.org/10.1029/2008JG000850
Дополнительные материалы
- скачать ESM.docx
- Приложение 1.
Таблица 1S. Статистические характеристики метеорологических показателей и их линейных временных трендов за весь период наблюдений 1998÷2022 гг. (n = 25).
Таблица 2S. Статистические характеристики Q10 and SR10 и их линейных временных трендов за весь период наблюдений 1998÷2022 гг. (n = 25).